Cell Biology Learning Track(I)膜结构
这个笔记主要基于小编老师的PPT的顺序进行总结的,所以就直接从细胞质膜开始了,对于前面细胞概述的内容咱们就直接略过了,这一块大家应该都有了解。
细胞质膜的理论模型的构建过程
(这一块其实是老生常谈的内容,小编简单说下,只是表达对教学内容的尊重,另外,本人的老师经常说细胞生物学是实验科学,所以任何一项的发现都是以实验作为支撑的,所以小编也谨记于心,对于以后任何一个理论的提出和讲述都会提到背后的实验依据~)
(但是对于做出这些工作的前辈科学家们,可能不会提及全名,但是发自内心地敬重)
1. 首先科学家们发现细胞膜对于脂溶性物质的通透性明显高于水溶性物质,那么因为相似相溶的原理呢,他们就猜测,细胞膜是脂肪栅(也就是有脂质分子组成的结构)
2.然后呢在这基础上再加上对于哺乳动物红细胞的观察(没有细胞器),科学家就裂解了红细胞提取脂质分子,然后将脂质分子展成单层,计算红细胞表面积(利用显微观察+立体几何知识)和脂质分子铺展的面积,大概得出脂质分子铺展开的面积是红细胞表面积的两倍,那么就推测这个膜结构的脂质是双层排列的
3. 接着发现细胞膜的张力小于纯脂质膜的张力,所以推测细胞膜上存在蛋白质
但是关键就来了,蛋白质是怎么分布在细胞膜上的呢?最初因为在电镜下观察到细胞膜呈现一个暗亮暗的三级结构,由于分子质量小的应该对电子吸收少一些,所以呈现亮,所以认为质膜是三明治结构即蛋白质-脂质-蛋白质,但是这个理论解释不了很多东西,但是莫名其妙地存在了很多年没有被推翻
解释不了的现象比如膜应该具有流动性的,但是这种单位膜模型(内核就是三明治模型)认为质膜是静态的,同时如果蛋白是简单覆盖在膜上的,那为什么我们很难提取膜蛋白?同时又如何解释细胞膜的厚度不均匀。
那么最后提出了流动镶嵌模型,认为蛋白质可以贯穿于脂质层,也可以结合在脂质层的两侧,脂质分子和蛋白质分子都是具有流动性的。
之后大家可能听说过脂筏模型,其实这个模型说的基本理论还是流动镶嵌模型,只是在对膜结构有更加清楚的认识之后,我们进行了细化。后面我们对脂质分子有了解之后,我们再提这个模型(因为小编当初在没有了解后面内容看这个模型也是似懂非懂)
但是有一句话大家得记着:生物膜可以看成是蛋白质在双层脂分子中的二维溶液
这里小编先谈一下自己对这句话的理解,首先什么是二维溶液?我们知道我们平常说的溶液一般我们的量度是体积,那么这个溶液就相当于三维的,这里提及的二维溶液呢主要是因为我们的膜结构就是由双层脂分子形成的,不管是溶剂分子(脂质)还是溶质分子(蛋白质)在这个平面结构当中都具有较高的自由度,所以我们认为这是一个溶液,那么这个溶液同时也对应了胞质和胞外的水溶液环境。但是需要说的是,这个二维溶液认为是一个形象化说法是没有问题的,但是这个比方不能认为溶质和溶剂的移动是不受拘束的,比方说细胞之间的黏连蛋白会使得膜的流动间断。
质膜的基本成分
首先要声明的是,这里涉及到的不是某一种分子,而是具有统一结构模式的某一类分子,这个类体现在两个方面(对于甘油磷脂和鞘脂而言):1. 头部的磷酸基团连接的基团是什么 2. 尾部的脂肪链(对于鞘脂还有烃基链)的组成是什么
1. 甘油磷脂(Glycerophosphatide)
甘油磷脂,或者简称磷脂,是膜脂的基本成分,占整个膜脂的50%以上,基本的结构如下
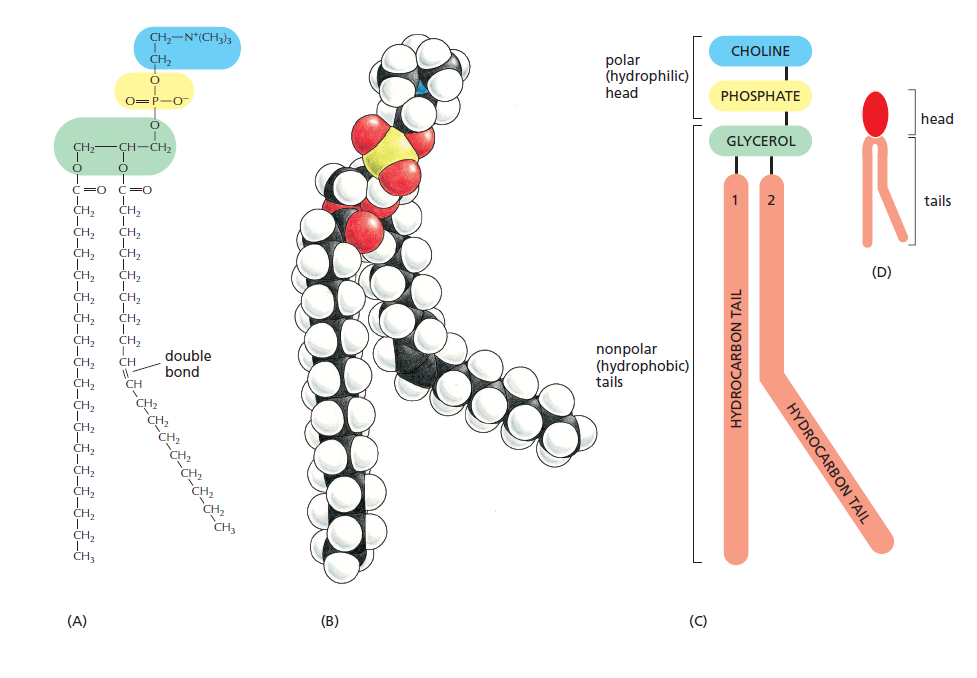
图片来源:Molecular Biology of the Cell (Alberts)(后续不再提及)
这个是以卵磷脂为例,不是所有的磷脂都是以胆碱作为头部的
那么可以看出磷脂分子可以看做有四个组成部分,基本骨架是甘油,其中甘油两个相邻的羟基和两分子脂肪酸发生酯化反应,连接两个脂肪酸链,那么一般来说有一个脂肪酸链是饱和的,一个是非饱和的,而且这个非饱和键是顺式的(向外拐),虽然图上只画了一个双键,但是可以存在多个双键的,甘油剩下的一个羟基和磷酸基团脱水结合,磷酸基团再结合一个亲水基团比如这里的胆碱
我们可以从结构中看出,磷酸分子是有一个亲水的头部和疏水的尾部的,同时可以稍微注意脂肪酸链的碳原子数都是偶数(因为合成脂肪酸链的时候是两个两个碳原子一连的)
那么磷脂有以下几种常见的类型,这些磷脂都参与形成膜结构,同时膜结构的不对成性的原因之一就是膜脂分子排布的不对称性
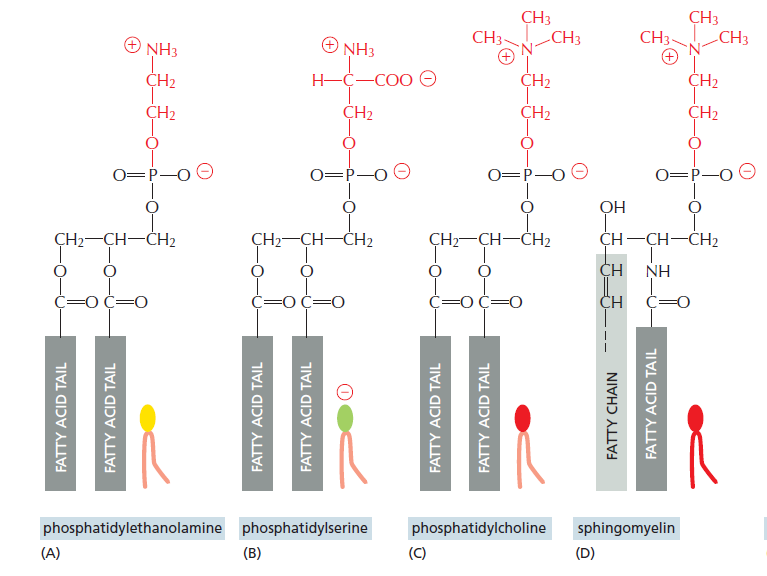
(A)磷脂酰乙醇胺(PE)(脑磷脂),主要特点是其磷酸基团连接的是一个较小的亲水基团乙醇胺,那么在构建磷脂双分子层的时候,正式由于其头部小,所以经常出现在膜拐角的内侧
(B)磷脂酰丝氨酸(PS)这个主要关注到丝氨酸是电中性,所以整个PS分子就带上了负电,那么膜电位的内负外正一部分就是由此贡献的,(由此推测这个磷脂多分布在膜内侧),那么如果PS发生了flip-flop,那么就可能介导细胞凋亡
(C)磷脂酰胆碱(卵磷脂)(PC),主要特点是磷酸基团连接的事一个较大的亲水基团胆碱,那么这个和(A)对应出现在膜拐角处的外侧
除此之外,还有一种磷脂称为磷脂酰肌醇(PI),磷酸基团连接有肌醇分子,肌醇分子是一个刚性的六元环结构,并存在5个羟基,可以作为氢键的供体与受体,由此可以与膜蛋白进行非共价结合和介导信号传导(有活化基团)
那么这里特别关照一下卵磷脂,人体所需的外源性胆碱90%是由卵磷脂提供的,由于重要神经递质乙酰胆碱的前体之一就是胆碱,所以卵磷脂的摄入对维持神经系统的正常功能至关重要,同时卵磷脂也可以降低胆固醇在血液中的含量
2. 鞘脂(sphingosine)
鞘脂包括鞘磷脂和鞘糖脂两类,那么我们以鞘磷脂为例说明鞘脂基本结构(D)
鞘脂不同于磷脂,其基本骨架为鞘氨醇,一个碳原子连接烃基(长度可变),一个碳原子连接亚氨基通过肽键连接脂肪酸链,最后一个碳原子醚键连接磷酸基团,磷酸基团再连接一个亲水基团(这里是胆碱)
那么鞘糖脂与鞘磷脂的不同在于最后一个碳原子不再连接磷酸基团,而是连接糖分子(含量约占膜脂5%,比例还是很大的)
我们再具体看看鞘磷脂和磷脂的差别,除了基本骨架的不同(鞘磷脂还露有一个羟基),鞘磷脂的脂肪酸链的长度一般是大于磷脂分子的,所以有鞘磷脂分子构建的脂双层的厚度是大于磷脂分子的
3. 胆固醇(Cholesterol)(固醇的代表,我们这里直接看胆固醇结构)
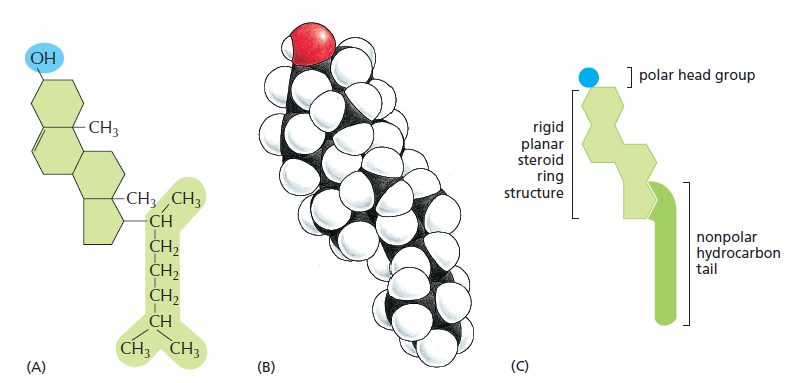
(初看这个结构就感觉这分子姿势很殷勤,功能肯定不简单)
胆固醇的结构可以分为三个部分,一个是小小的羟基作为亲水的头,刚性的环结构,还有疏水的烃链,胆固醇主要分布在动物细胞膜上,极少分布在原核生物膜。
由于胆固醇侧向的结构,使得其不能独自形成双分子层,多插入甘油磷脂分子之间(因为鞘脂的尾部更加紧密)。那么在正常的生理温度下,因为胆固醇分子实际上间隔磷脂分子,刚性环结构实际上也使得膜结构也更加稳定(但是对流动性影响不大),如果温度下降,那么胆固醇就可以阻碍磷脂分子的聚集,使得分子之间的作用较弱,所以当温度更低的时候,磷脂分子之间的作用才会克服热运动使得膜结构凝固,所以在此温度下,胆固醇的存在实际上有利于膜流动性的维持。
那么既然说到这里,我们接着谈膜流动性是如何在不同环境下维持的。
我们知道膜的流动性的适中对于生命活动是非常重要的,如果流动性太低太高物质的运输、信号转导都会受到影响。那么很多变温动物需要维持其膜的流动性在一定的范围内,这一点可以通过合成链更短的脂分子(分子间作用更小,在低温下流动性更好)、增加不饱和键的个数(不断形成kink,原理同上),或者插入更多的胆固醇等
(如果膜鞘磷脂的含量过高,那么就会降低膜的流动性,比如动脉粥样硬化病变膜脂就高于正常的动脉组织)
膜蛋白
膜蛋白根据其所处的膜的位置,可以分为三类,一类是分布在膜两侧的称为周边膜蛋白(peripheral membrane protein)或者是外在膜蛋白(extrinsic membrane protein),整合膜蛋白(integral membrane protein)或内在膜蛋白(intrinsic membrane protein),脂锚定膜蛋白(lipid-anchored membrane protein)
那么跨膜蛋白的跨膜区段应该是疏水基团为主,在内部多是亲水基团。那么由于膜的流动性,可能在蛋白质最初运输到膜上的时候并没有结合,但是在这二维溶液的流动下就可以结合形成蛋白复合体。
(这里插一句,可以利用一种分子(一端亲水一端疏水)作为去垢剂(detergent)来分离膜蛋白)
膜流动性
在胆固醇那里提及的不再赘述
膜蛋白流动性的验证可以通过荧光标记的细胞诱导融合,也可以用抗体结合膜上高度表达的蛋白,观察是否出现成斑或者成帽现象
膜蛋白流动性的定量测定也可以利用荧光漂白回复(FPR)技术,首先用荧光标记膜蛋白,使得被标记膜蛋白在细胞膜上分布均匀后用激光猝灭某一区域的荧光,然后检测该处荧光强度的恢复速率即可求出膜蛋白流动速率
膜不对称性
膜不对称性主要体现在三个部分
1. 膜蛋白分布的不对称性,由于膜蛋白功能的不同,如果是作为外界信号受体蛋白,那么一般分布在膜外侧,如果是作为内部结构稳固的连接蛋白,那么一般在膜内侧(如血影蛋白)
2. 膜糖蛋白、糖脂分布的不对称性:糖基全部分布在膜外侧
3. 膜脂分布的不对称性:这一点在介绍膜脂的时候已经说明了
(但是胆固醇分布是相对对称的)
那么膜的不对称性就会使得膜的功能具有方向性,进而促进生命活动的方向性和有序性
那么最后来提到脂筏模型(还好没忘记)
其实根据我们对于膜结构以及膜分子的了解之后,我们可以想见这样一个事实,在细胞膜上可能会存在某种功能分区,就如同我们在三维细胞胞质当中有细胞器一样,这里可能存在着某种以膜脂为介质的特定种类蛋白集中分布相对固定的分区,这个分区同时也随着膜的流动性一起流动,那么这就是脂筏模型,那么这个分区的维持可能就需要尾链更长的鞘磷脂作为中坚力量,同时插入的胆固醇的含量也可能增多来使得这个分区的结构是相对固定的
那么简单总结一下,我们通过膜的分子结构的分析,可以理解膜的不对称性和膜的流动性,为我们之后学习物质运输奠定了坚实基础
(另外此章小编的老师还讲了红细胞质膜的特点,我觉得和前面不太连贯,这里提下只是提醒小编记得复习哈哈~~)