神经流形:大脑功能几何基础的革命性视角
论文基本信息
标题:A neural manifold view of the brain
期刊:Nature Neuroscience(影响因子28.5,神经科学顶级期刊)
文章类型:综述文章(Review Article)
接收时间:2023年7月21日提交,2025年6月30日接受,2025年7月28日在线发表
作者单位:蒙特利尔大学神经科学系、Mila-魁北克人工智能研究所、伊拉斯谟医学中心神经科学系、帝国理工学院生物工程系
通讯作者:Matthew G. Perich和Juan A. Gallego
文献编号:Nature Neuroscience Volume 28, August 2025, 1582-1597
DOI:https://doi.org/10.1038/s41593-025-02031-z
引言:从神经元到流形的范式转变
这篇发表于Nature Neuroscience的重要综述,标志着神经科学研究范式的重要转折。传统神经科学基于"神经元学说",将单个神经元视为大脑的基本计算单元,通过将单个神经元活动与实验者定义的任务变量相关联来理解脑功能。然而,动物行为最终产生于神经群体的复杂协调,这使得从单个神经元视角理解脑与行为的基本关系变得困难。
随着现代实验技术能够同时记录大量神经元活动,神经科学的焦点已转向研究神经群体的协调活动。跨越多物种、脑区和行为的实证结果一致表明,神经群体活动可以被相对较少的全脑活动模式所捕获,这一看似令人惊讶的发现引出了低维潜在动力学存在的假设。本文提出,这些经验观察反映了低维潜在动态的存在,这些动态源于神经流形对神经元群体活动的约束。
神经流形的数学本质与精确定义
流形的基本数学概念
在数学上,流形被定义为局部欧几里得拓扑空间。许多类型的连续几何对象,如平面、环面或球体,都符合这一定义。例如,球体或咖啡杯的表面可以被认为是物理世界三维空间中的二维流形。
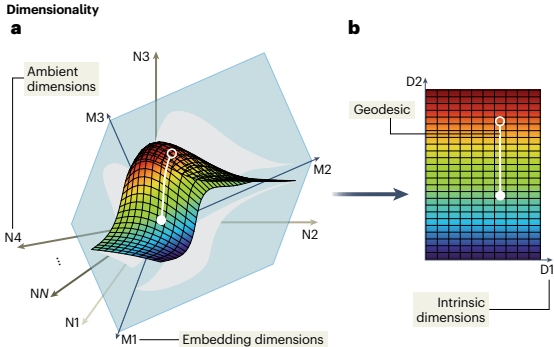
在现有神经科学文献中,"流形"一词具有多种含义,包括:(i) 降维或流形学习方法的输出,旨在可视化或探索数据,或用于脑机接口等工程应用;(ii) 神经群体活动的几何组织, presumably 沿神经状态空间中的表面;(iii) 在特定行为期间捕获神经群体活动随时间变化的低维结构。
神经流形的特定定义
本文采用以下特定的神经流形定义:捕获神经群体动力学在考虑其生物物理约束下可以遍历的可能集体状态的低维表面。因此,我们认为神经流形对大脑中的神经活动提供了有意义的约束。
虽然神经流形通常使用降维或流形学习方法从神经群体记录中估计,但仅应用这些方法并不必然意味着神经流形作为对神经群体活动的有意义约束而存在。神经流形包含几何和动态特征,因为这两个方面紧密相连。
维度概念的精细化
论文中精确定义了三种维度概念:
内在维度:完全参数化流形结构所需的独立变量数量
嵌入维度:完全包含流形所需的正交状态空间方向的最小数量
环境维度:可用于估计神经流形的轴数,在神经科学实验中通常是记录神经元的数量
这种分类法没有明确定义"低维"和"高维"的概念,这些概念必须相对于记录神经元的数量或行为的维度等特征来定义。由于共同输入、循环连接性和动物行为等众多约束,我们预期底层神经流形的内在维度将远少于记录神经元的数量。
神经流形的实验推导与验证方法
神经状态空间的构建
神经流形在神经状态空间中进行估计,其中每个正交轴代表数据集中单个记录神经元(或独立多单元)的放电率。在数据集的每个时间步,神经群体活动可以表示为该状态空间中的一个点。随着时间的推移,这些点的集合通过状态空间追踪一条潜在轨迹。
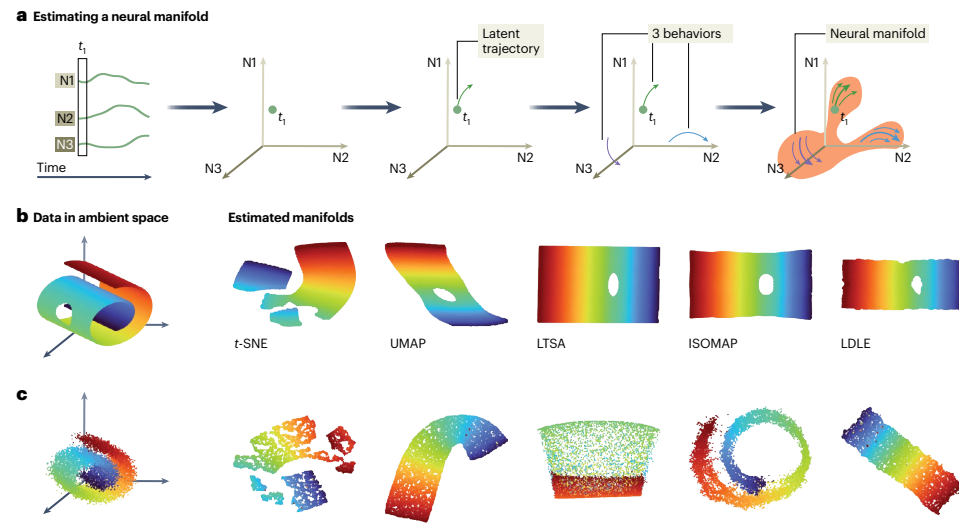
在此框架中,给定群体中每个神经元的活动可以被视为这条潜在轨迹在定义神经状态空间的一个轴上的(随机)投影。神经流形框架提出,数据集中所有时间点的潜在轨迹集合将探索可能神经状态的受限子集,产生内在低维结构,我们假设这种结构被限制在神经流形的表面上。
降维与流形学习方法
神经群体的协调活动可以通过相对较少的群体活动模式很好地捕获,这是由于组成神经元放电率的相关性和冗余性。能够识别此类低维活动模式的工具被称为降维方法。
线性降维方法(如PCA或因子分析)将原始数据描述为潜在信号的线性组合。这些潜在信号捕获了神经元间的协变模式(即神经状态空间中的方向),从而定义了底层神经活动的超平面——一个平坦流形。在神经流形是非线性的情况下,这些线性方法最多只能提供对真实底层流形的平面近似,导致估计的维度高于真实内在维度数。
非线性流形学习方法可以直接纳入关于流形的假设(如连续性或几何),从而提供对底层群体结构的更好估计。例如,Isomap通过保持嵌入中相邻点之间的测地距离来构建保持原始环境空间整体内在几何的低维嵌入。其他方法包括局部线性嵌入、扩散映射、拉普拉斯特征映射等。
每种流形估计方法都有其自身的假设和优势,技术选择会极大地影响嵌入结果。例如,使用PCA等线性方法估计的平坦流形,如果研究者没有选择足够的维度,可能会错过重要的拓扑特征,因为行为相关特征可能在高方差成分中被捕获。
全脑范围内的神经流形证据
跨物种的一致性发现
研究表明,具有行为相关组织的低维潜在动态的存在与神经群体活动受到底层神经流形约束的概念一致。因此,任何可能的行为状态都与沿着流形的神经状态相关联。
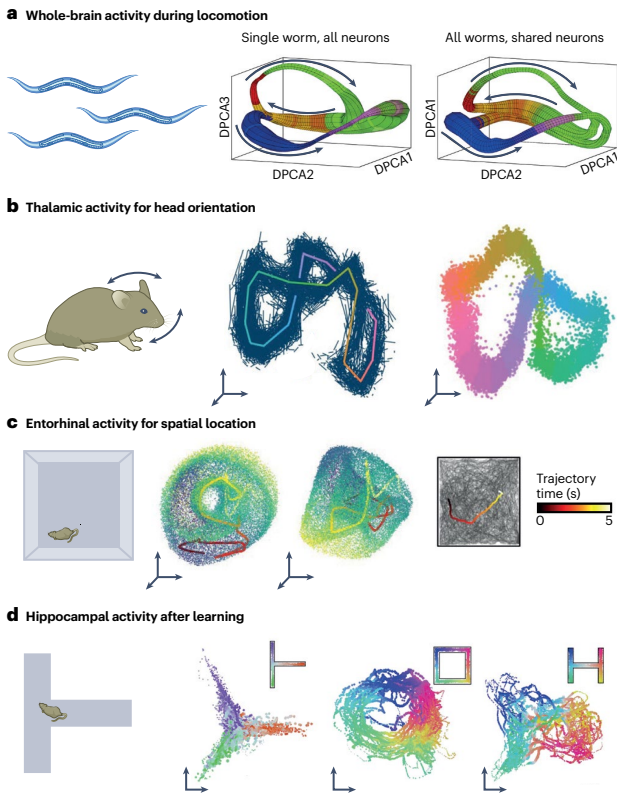
近期工作直接识别了底层神经流形:(i) 在线虫神经系统所有神经元状态中识别了与 locomotion 期间一致的低维几何结构,这些神经流形的几何结构在不同个体间高度保持,流形内位置与运动行为之间的映射也是如此;(ii) 在不同小鼠丘脑头部方向细胞中观察到一致的神经流形拓扑结构:神经群体动力学可以通过不规则环形吸引子流形描述,这种几何组织在清醒和睡眠状态间持续存在;(iii) 内嗅皮层网格细胞群体在不同环境导航期间显示出保持的环面拓扑结构,在睡眠期间也持续存在;(iv) 证据积累沿小鼠海马体中也表示在连续非线性流形上,该流形也表示空间;(v) 攻击状态与下丘脑中的线吸引子流形相关联;(vi) 非线性流形被证明能最好地捕获小鼠、猴子和人类多个皮层和下皮层运动区域与运动相关的活动。
流形属性与行为功能的关联
神经流形的属性可以与行为功能和执行的任务的特征密切相关。至少在某些脑区,神经流形是由记录神经群体的行为功能塑造的。确实,小鼠丘脑和斑马鱼前后脑中的环形吸引子流形允许准确跟踪头部方向,小鼠内嗅皮层中发现的环面流形结构能够构建空间的准确表示。
海马体空间"地图"也利用流形属性进行导航:随着小鼠熟悉新环境,其海马体CA1流形中的活动越来越反映环境的形状。同样,流形属性在塑造行为功能方面可以发挥重要作用。在内侧前额叶皮层中,沿假定流形的轨迹组织匹配猴子定时内部估计。此外,任务相关潜在动态表现出弯曲几何形状——可能由非线性流形产生——使得这种曲率的线性读出解释了猴子在估计时间间隔时的响应模式。
神经流形的形成因素与多时间尺度演化
内在生物物理约束
神经流形可能从网络中的内在约束中出现。首先,主导群体结构在麻醉猫和清醒小鼠的视觉皮层自发和刺激诱发活动间保持保守,小鼠丘脑头部方向细胞和内侧内嗅皮层网格细胞底层神经流形的拓扑结构在行为和睡眠间持续存在。其次,小鼠视觉皮层中足够多刺激辨别神经元的全息光遗传激活重现了同一区域不同神经元群体在刺激诱发活动期间描述的潜在轨迹。
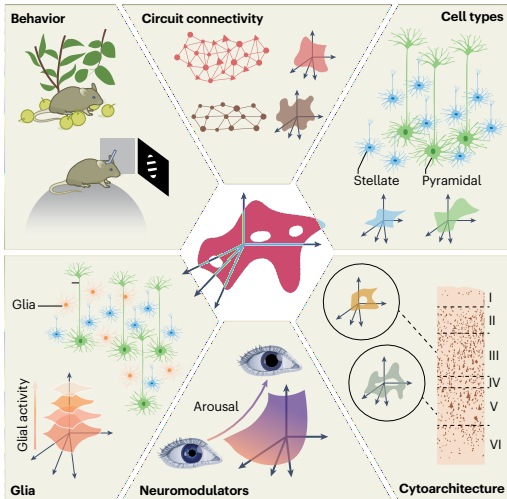
神经流形应由电路组织方面(如突触连接、细胞类型、有效突触强度)塑造。基因指定特性、输入和投射目标应约束神经群体活动,从而约束神经流形的出现。在更高层次上,电路中的连接性(如连接 motif、层状组织)应类似地塑造和约束群体活动。确实,不同脑区——已知具有不同的连接谱、细胞组成和输入——在估计流形中表现出不同程度的非线性、潜在轨迹的独特结构和群体活动中变化的维度。
行为与任务的塑造作用
行为以两种关键方式塑造观察到的神经流形属性。首先,神经流形的内在维度可以反映动物在记录期间探索的行为或认知变量的数量。对于前额叶皮层,这可能是探索期间探索的选择范围,而对于运动皮层,可能是动物执行的不同运动。神经记录通常在实验室环境中执行任务期间进行,因此许多实验室任务中感觉刺激和/或行为输出的相对低维度约束了神经群体访问的状态范围,这可能有助于观察到的神经流形的低内在维度。
其次,如果任务或行为输出中存在几何或结构,我们应期望在神经流形的某些方面看到这种结构的反映。例如,运动任务中到达目标的空间组织可能确保估计的神经流形中存在类似组织。
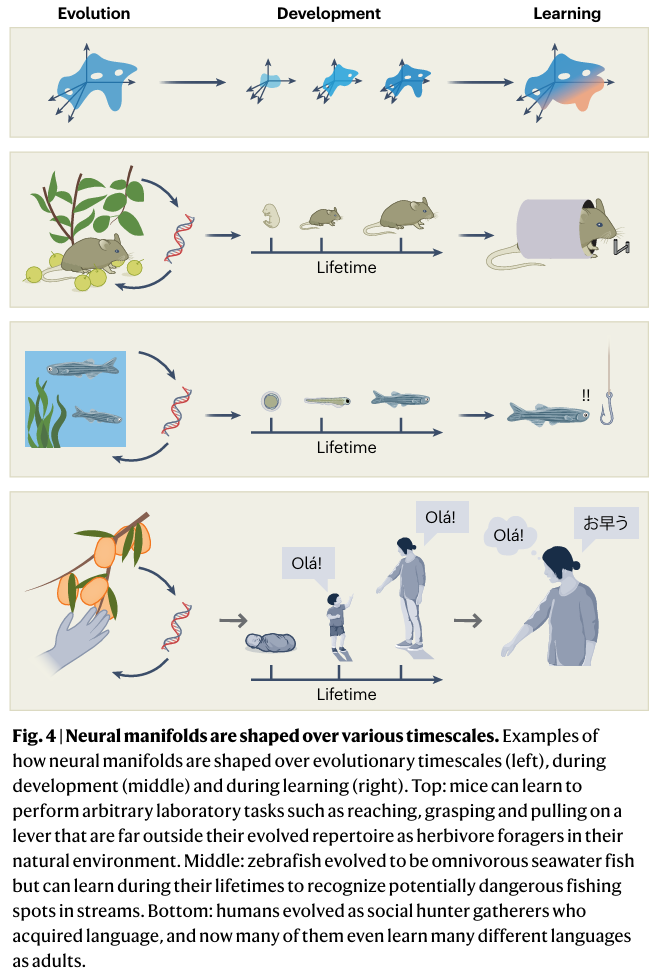
多时间尺度的演化
神经流形及其属性在多个较长时间尺度上同时塑造:学习、发育和进化。相同行为的一致执行似乎与同样稳定的流形相关联,假设所有其他内部状态(如冲动性、参与度)等效。然而,这种表面稳定性并不意味着神经流形是不可变的。在学习时间尺度上,神经流形属性可能改变以帮助通过练习提高执行某些行为的能力,或获得非本能行为。
神经流形在行为和学习时间尺度上出现,受到发育和进化更长的时间尺度约束。在最长进化时间尺度上,动物获得适应,然后成为物种本能行为 repertoire 的一部分,以及它们的学习能力。这些适应在发育时间尺度上通过运行"遗传程序"表达,最终产生具有其物种基本神经回路特性的个体。这些回路特性有助于约束大脑在发育本能行为和获得新的学习行为时产生的可能活动模式,这一过程可能对形成所需的神经流形是必要的。
因果操纵与实验验证框架
三类因果实验目标
发展脑功能的多元解释需要因果操纵,使我们能够直接验证、表征和操纵神经流形,超越通过神经记录的观察。我们提出因果实验的三个总体目标:
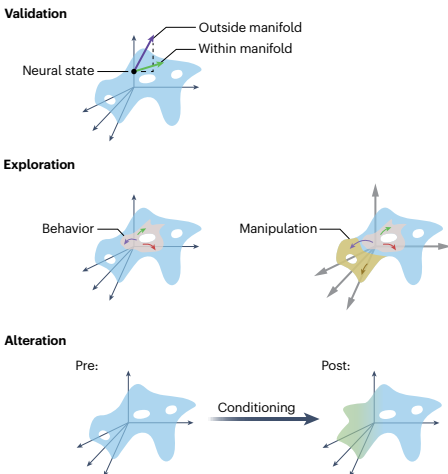
验证:实验者因果测试神经流形是否以及如何约束神经和行为状态
探索:实验者探查现有流形内可能的神经状态,以及该流形如何约束未来或未观察到的活动
改变:实验者修改现有流形以研究它如何塑造动物行为(反之亦然)
因果操纵以表征神经流形有两个关键属性。首先是扰动的时间尺度;短暂操纵通常对神经回路和行为有不同的影响,而不是长期、延长操纵。其次是扰动效果是否将神经状态带到流形上或流形外。流形上扰动是将神经活动移动到动物行为期间可以达到的状态。如果我们假设神经流形状态和行为状态之间的关系保持,流形上扰动的预期结果是动物最终将产生或经历预期的行为状态。
理论意义与未来发展方向
神经流形的涌现特性
神经流形提供了概念化神经群体协调活动的直观方式,使得能够整合跨研究结果、比较跨脑区,并基于抽象神经电路实现细节的几何和动力学进行可检验的实验预测。一个重要问题是神经流形描述层次是否比其低层次组成部分(如电路、神经元和突触)具有更多解释力。
在这种情况下,神经流形将被视为涌现的。为说明这一点,我们做一个类比:如果你的朋友问你关于刚看过的电影,你不会通过仔细描述电影中的一百帧来解释其情节,而是总结关键事件。类似地,虽然神经流形根植于生物物理特性,但它捕获了神经群体活动中直接告知动物行为的结构。因此,它可能提供比从其组成神经元、突触等得出的更紧凑和可理解的大脑功能描述。
全脑整合与多尺度组织
神经流形及其相应的潜在动态历史上在单个区域(或可能两三个小集合)内识别。然而,产生行为的神经活动是众多高度专业化区域之间连续相互作用的产物。因此,在将神经流形属性与行为关联时存在一个关键的开放问题:当我们在小(如皮层柱)、中(如区域)和大(如全脑)尺度之间移动时,应考虑大脑中流形的空间尺度是多少?
一种可能性是大脑中的每个子模块(细胞类型、皮层柱、区域)具有其自己的独立流形和动力学,由其局部电路特性和行为特定输入决定。然后,假定的"全脑流形"可以通过组装这些区域特定流形来完全描述。然而,我们认为这不太可能,考虑到这些子模块组织中的高度互连性。说明问题,我们考虑由给定皮层区域中所有神经元组成的群体。这些神经元可以进一步细分为皮层柱。现在考虑仅这些柱中一个底层所有神经元活动的神经流形。该流形反映了组成神经群体的协调活动—— presumably 在某种程度上归因于可能状态的约束。这种协方差(或约束)可能源于该群体内的局部(循环)连接,或来自驱动神经元的输入,例如来自其他附近柱。因此,每个柱特定流形是其他柱输入的产品,每个柱具有其自己的柱特定流形。我们可能无法表征底层区域所有神经元活动的总体流形,而不考虑所有柱之间的相互作用。
循环神经网络的理论启示
循环神经网络(RNN)为研究神经流形提供了探索和生成关于神经流形与行为关系的假设的平台。RNN是通过循环连接单元构建的函数逼近器,这些单元可视为类似于生物神经元。这些连接然后使用数学技术进行训练以实现某些目标。
典型RNN的动力学由以下公式控制:
在这里,每个第i个单元的动力学x˙i(t)受三个组成部分影响:(i) 其自身状态xi(t);(ii) 来自其他单元xj(t)的输入,非线性变换并由Jij加权;(iii) 外部输入u(t),其中第k个输入影响第i个单元加权由Bik。许多RNN通过非线性激活函数ϕ对循环权重施加非线性,并且可能包括随机项。
连接矩阵J中的结构影响RNN群体活动的功能、动力学和维度。该结构中与神经流形相关的关键组成部分是"秩",它指的是重建矩阵所需的最小正交向量数l。如果l∼N,则N×N连接矩阵被认为是"满秩",如果l<<N,则是"低秩"。结构化连接使研究人员能够研究大脑中低维动力学的出现。
结论:迈向统一大脑理论的几何基础
神经流形框架提供了比其组成神经元的独立活动更紧凑且通常更可解释的描述神经群体功能的方法。这一框架可以导致生成关于脑功能的清晰可检验假设。此外,神经流形框架还可以增强我们利用这些见解治疗神经系统疾病和开发脑接口新神经技术的能力。
最终,神经流形框架可以帮助连接行为神经基础的所有方面——认知、行动、感觉和情绪——以实现大脑工作方式的统一描述。随着新技术允许我们记录和操纵更大神经群体,以及计算方法的进步使我们能够估计和解释更复杂流形,我们正站在神经科学新时代的门槛上,其中几何原理可能提供我们一直在寻找的统一语言。
这篇综述不仅总结了当前最新技术,而且为该领域的未来研究绘制了路线图,强调了需要理论、实验和计算模型结合来解决剩余挑战,如全脑流形如何组织、流形属性如何通过学习修改,以及神经流形框架是否可以提供治疗神经系统疾病的新途径。