项目文章|MeRIP-seq助力解析m6A RNA甲基化与康乃馨花衰老的调控机制
近日,华中农业大学果蔬园艺作物种质创新与利用全国重点实验室和湖北洪山实验室张帆教授课题组在The Plant Journal发表了题为“Dynamic m6A RNA methylation correlates with ethylene-induced petal senescence and may modulate antioxidant and metabolic pathways in carnation (Dianthus caryophyllus L.)”的研究论文。该研究发现m6A RNA甲基化修饰通过抗氧化和代谢途径调控乙烯诱导的康乃馨花衰老过程。

研究背景
花衰老是植物生命周期中的关键阶段,受乙烯等多种激素调控。尽管N6-甲基腺苷(m6A)RNA修饰在植物发育和逆境响应中具有重要作用,但其在乙烯诱导的花衰老中的功能尚不明确。康乃馨(Dianthus caryophyllus L.)作为乙烯敏感型切花,是研究花衰老的理想模型。本研究旨在探讨m6A修饰在乙烯信号下如何动态变化,并解析其通过抗氧化和代谢途径影响康乃馨花衰老的分子机制。
研究路线
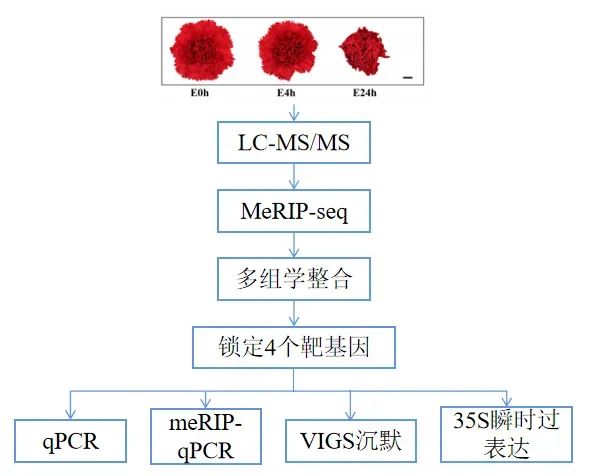
研究结果
1. 在乙烯诱导的康乃馨花衰老过程中,mRNA m6A水平呈现出动态变化
首先,使用LC-MS/MS技术分析了乙烯处理不同时间(0h、4h和24h)的康乃馨花中mRNA m6A的水平(Fig.1A)。结果发现,乙烯处理显著降低康乃馨花mRNA的m6A水平,mRNA中m6A/A的比例从E0h的0.58%降至E4h的0.47%,E4h与E24h之间没有显著差异(Fig.1B)。

Fig1A-B.乙烯处理下康乃馨花mRNA m6A甲基化图谱分析
2. 不同乙烯处理时间点下康乃馨花中的m6A特征
为了探究m6A修饰是否与乙烯诱导的康乃馨花衰老有关,进一步进行了MeRIP-seq(该部分由武汉爱基百客提供服务)。使用exomePeak分析软件来分析峰值信息,显著峰值的阈值设定为FDR<0.05。在所有三个生物重复样本中一致识别到的m6A peak峰为高置信度峰用于所有后续分析。在E0h、E4h和E24h下分别获得了4209个、5092个和4817个高置信度峰(Fig.1C),这些peak峰编码了2687个、3487个和3544个基因。接下来,计算了每个转录本上m6A峰的数量,有趣的是,随着乙烯处理时间的增加,含有两个或更多m6A峰的转录本百分比下降,从E0h的45.01%降至E4h的30.18%,再到E24h的38.77%,这表明大多数转录本以单个m6A峰的形式存在(Fig.1D)。
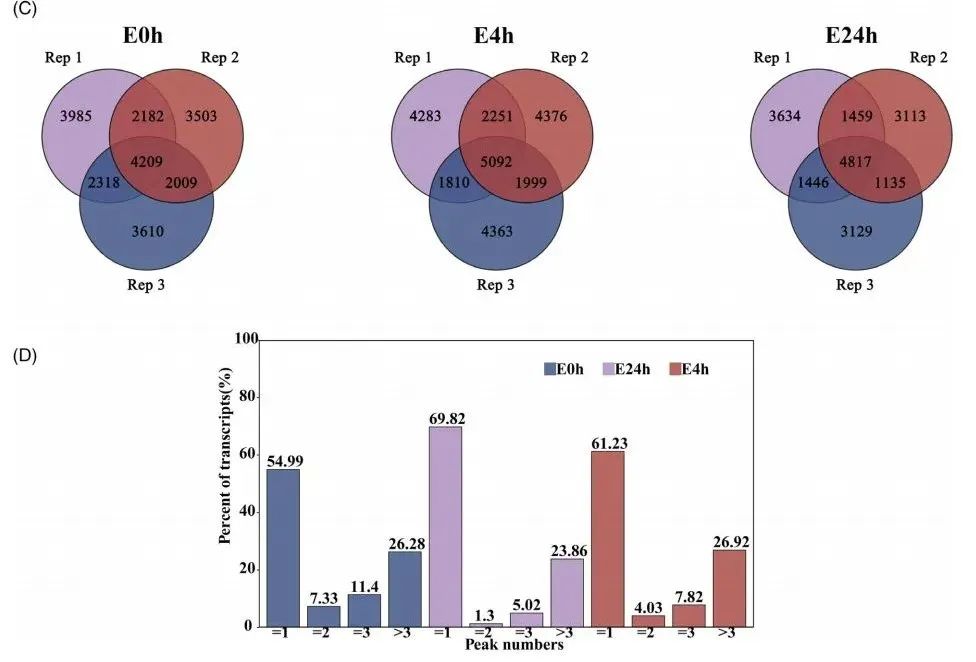
Fig1C-D.乙烯处理下康乃馨花mRNA m6A甲基化图谱分析
3. m6A分布在乙烯诱导的康乃馨花衰老过程中的变化
随后,作者评估了E0h、E4h和E24h三个时间点m6A峰的分布情况。在所有三个阶段中,m6A修饰主要富集于终止密码子区。值得注意的是,随着乙烯处理时间的延长,起始密码子区的m6A修饰呈下降趋势,而终止密码子区则显著上升(Fig.2A)。从m6A峰的绝对数量来看,CDS区占比最高,其次是终止密码子区,而5′UTR区最少(Fig.2B)。进一步分析发现,随乙烯处理时间延长,CDS区的m6A比例下降,而起始密码子区有所上升。对各基因区段长度进行归一化后,m6A修饰在所有三个时间点均主要富集于终止密码子及起始密码子邻近区域(Fig.2C)。这表明虽然m6A绝对丰度最高的是CDS区(Fig.2B),但按单位长度计算的修饰密度在终止密码子和起始密码子区更为显著。最后,利用HOMER软件鉴定到一个高可信度的保守序列基序:GVNGVN(Fig.2D)。
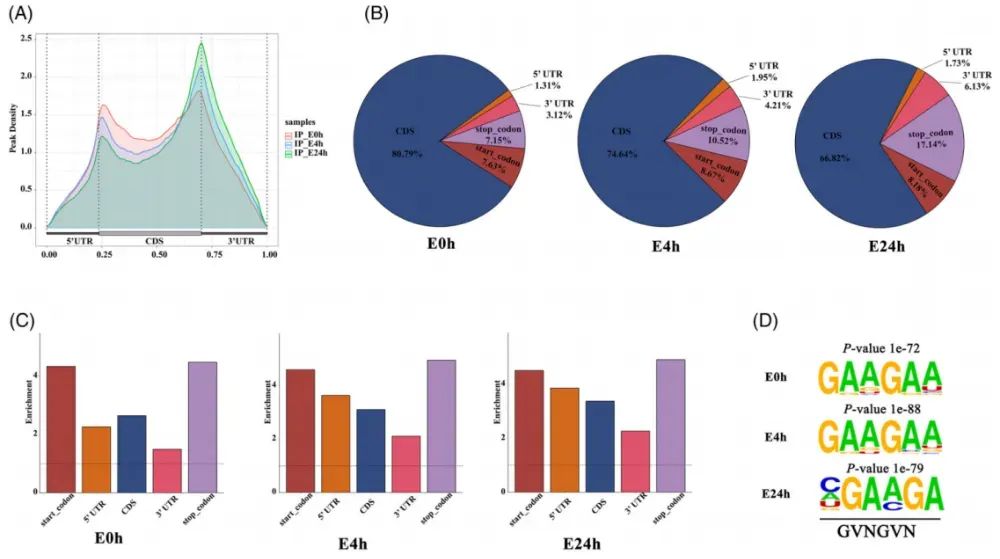
Fig2.乙烯处理康乃馨花过程中m⁶A的动态分布
4. m⁶A甲基化在整体上影响乙烯诱导的康乃馨花衰老过程中的mRNA丰度
为了弄清m6A重排是否伴随转录本丰度变化,作者把MeRIP-seq差异peak(|FC|≥1.5,FDR<0.05)与已发表的RNA-seq表达谱进行联合分析。使用累积分布曲线和箱型图来揭示基因表达比的整体趋势,分析表明:
E24h vs E0h:共1414个差异m6A峰(507个高甲基化、907个低甲基化)(Fig.3A),分别关联到473个和872个转录本(Fig.3B,3C);其中低甲基化基因整体表达下降,高甲基化基因表达上升(CDF曲线显著分离,Wilcoxon P<0.001)(Fig.3G)。
E4h vs E0h:770个差异峰(304高、466低)(Fig.3D),分别对应295个和444个转录本(Fig.3E,3F);表达变化趋势与E24h一致但幅度较小(Fig.3H),提示早期甲基化调控尚未完全放大。
KEGG富集显示E24h差异甲基化基因显著富集“核糖体”“TCA循环”“谷胱甘肽代谢”“氧化磷酸化”等衰老核心通路(Fig.3I)。这些结果表明m6A的动态变化与mRNA稳态水平正相关,且集中于能量与抗氧化代谢网络。
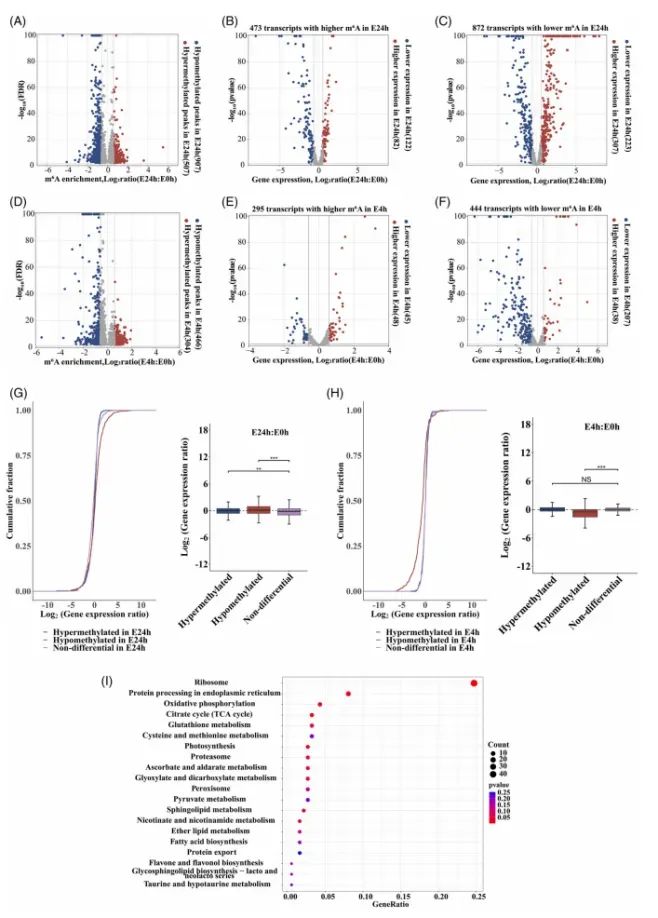
Fig3.乙烯处理的康乃馨花中m6A修饰与mRNA丰度之间的相关性
5. m6A修饰参与调节康乃馨中与乙烯诱导的衰老相关基因的表达
为了探究m6A修饰在康乃馨花中与乙烯诱导的衰老相关的基因中的作用,作者在GSH代谢及TCA循环通路中进一步筛选出4个同时满足“差异甲基化+差异表达”的基因,根据它们的功能注释和与拟南芥蛋白质的序列同源性,随后分别命名为DcGST10、DcGPX8、DcACLA和DcACLB。其中,DcGST10和DcGPX8是参与抗氧化系统的两个核心基因,在CDS区域显示出显著的m6A峰富集(Fig.4A,B)。从E0h到E24h,这些基因的m6A修饰水平和转录丰度均显著降低。qPCR和meRIP-qPCR实验进一步验证了m6A修饰与基因表达之间的正调控关系,表明m6A水平与转录丰度之间存在显著相关性(Fig.4C-E)。此外,与三羧酸循环相关的两个基因DcACLA和DcACLB表现出相似的趋势,在CDS区域有显著的m6A峰富集(Fig.4F,G)。进一步的qPCR和meRIP-qPCR分析证实,DcACLA和DcACLB的m6A甲基化水平与其从E0小时到E24小时的转录丰度呈正相关(Fig.4H-J)。这些结果表明,m6A以“增强mRNA稳定性/翻译效率”的方式正向调控这些基因。
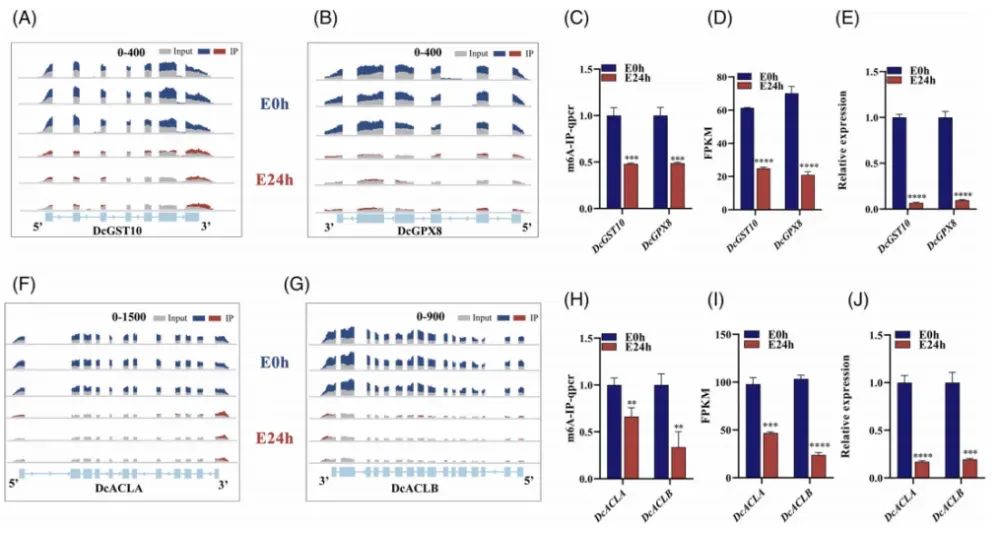
Fig4.在E0h和E24h时,康乃馨花中四个基因的m6A修饰和基因表达水平的比较
6. DcGST10和DcGPX8通过氧化应激和乙烯生物合成促进康乃馨花衰老
功能验证采用VIGS沉默(Fig.5)与35S瞬时过表达(Fig.6)双策略:
VIGS:DcGST10或DcGPX8表达量下降70–80%;瓶插寿命缩短3–4d;GSH含量下降约35%;DAB/NBT染色表明H2O2和O2-大量积累;乙烯合成基因DcACS1、DcACO1及衰老标记DcSAG12显著上调(Fig.5)。
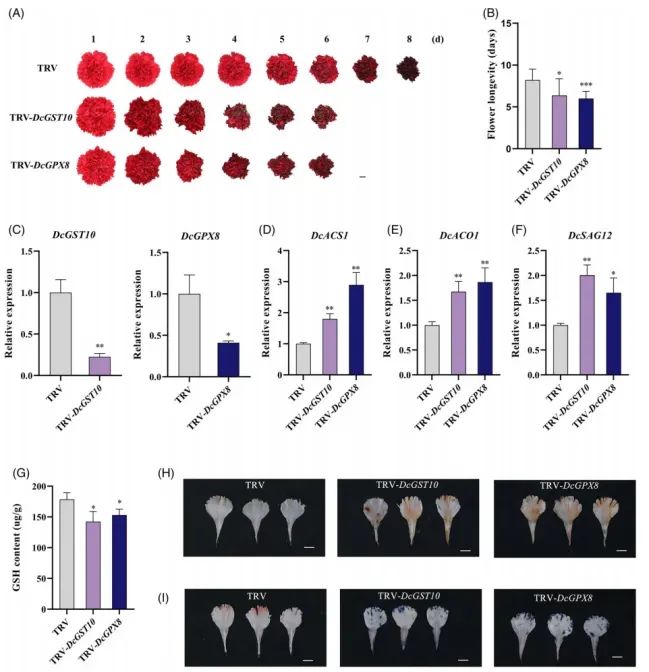
Fig5.DcGST10和DcGPX8的沉默加速了康乃馨花的衰老
过表达:DcGST10/GPX8表达量提高3–5倍;瓶插寿命延长4–5d;GSH水平升高50%;ROS减少;乙烯级联基因被抑制(Fig.6)。
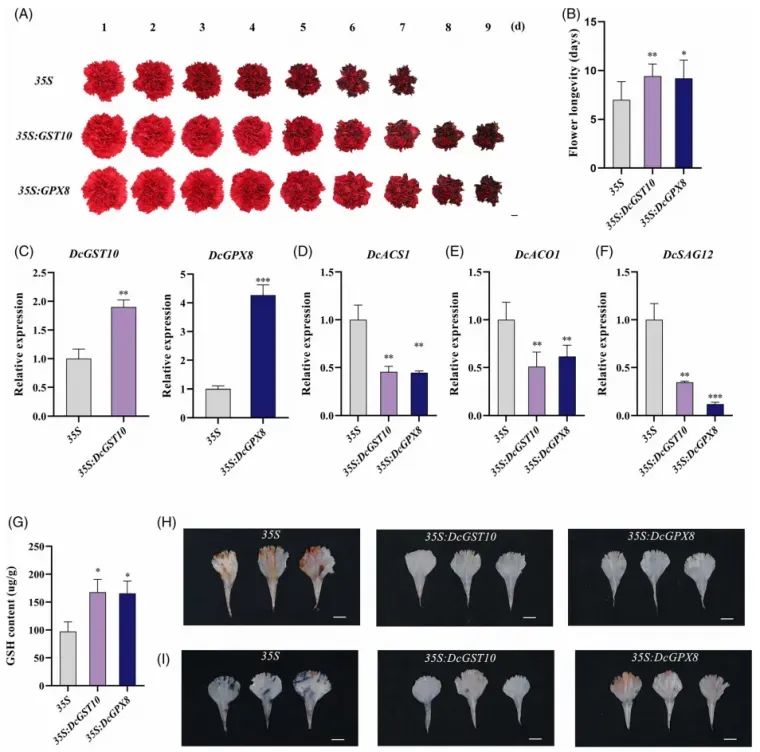
Fig6.过表达DcGST10和DcGPX8延缓了康乃馨花的衰老
花瓣圆片实验同样验证:沉默系褪色快、离子渗漏高;过表达系保绿、保色、膜完整性增强。综上,DcGST10/GPX8通过提升抗氧化能力抑制ROS的产生,从而延缓花衰老。
7. DcACLA与DcACLB通过清除ROS促进康乃馨花衰老
对DcACLA/B进行类似验证:
VIGS:ACL酶活下降约50%,瓶插寿命缩短2.5–3d;ROS显著积累;乙烯合成与衰老基因上调(Fig.7)。
过表达:ACL酶活提高60%,寿命延长3d;ROS水平降低;膜稳定性增强(Fig.8)。
DAB/NBT染色、离子渗漏结果与DcGST10/GPX8实验高度一致。
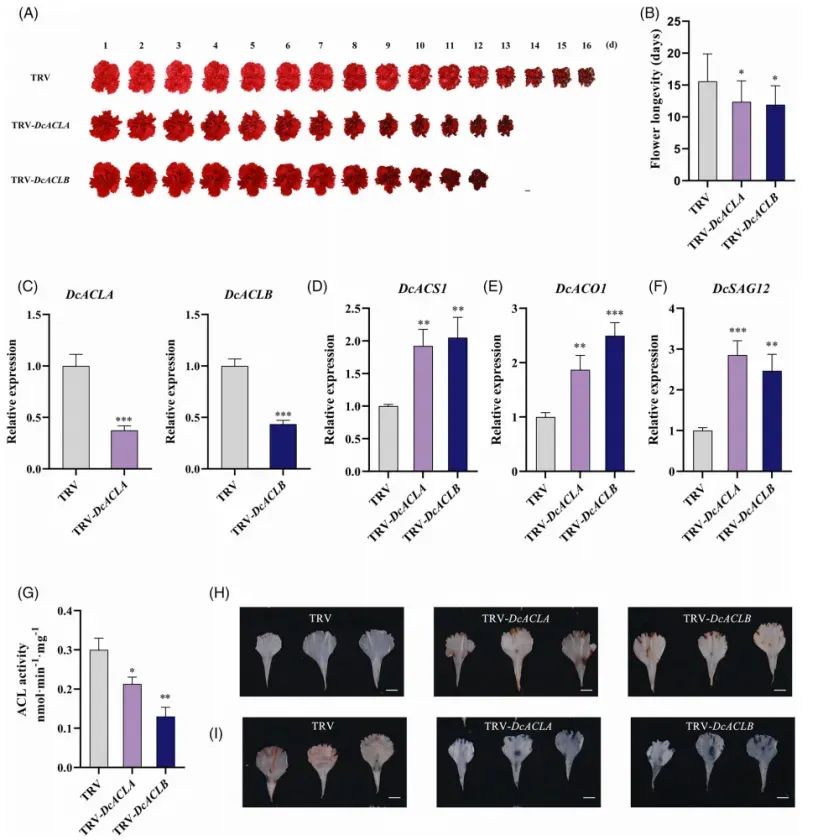
Fig7.DcACLA和DcACLB的沉默加速了康乃馨花衰老
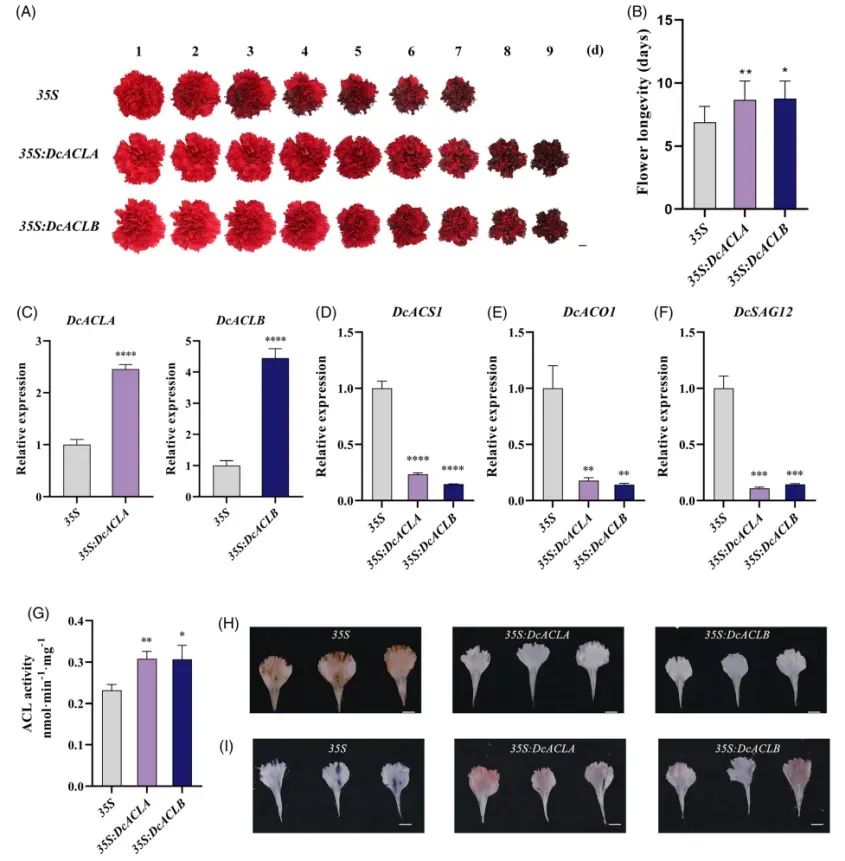
Fig8.过表达DcACLA和DcACLB延缓了康乃馨花衰老
总 结
本文研究了乙烯诱导的康乃馨(Dianthus caryophyllus L.)花衰老过程中RNA N6-甲基腺苷(m6A)修饰的动态变化及其分子机制。研究发现乙烯处理显著降低了花中m6A的整体水平,并通过MeRIP-seq分析揭示了m6A修饰分布模式的改变,特别是在终止密码子区域的富集增加。研究鉴定了四个与衰老相关的基因(DcGST10、DcGPX8、DcACLA和DcACLB),这些基因的m6A水平与其mRNA表达水平呈正相关。通过基因沉默和过表达实验,证实这些基因通过调节抗氧化和代谢途径影响花衰老进程。该研究为理解m6A修饰在植物衰老中的表观遗传调控网络提供了新的见解。
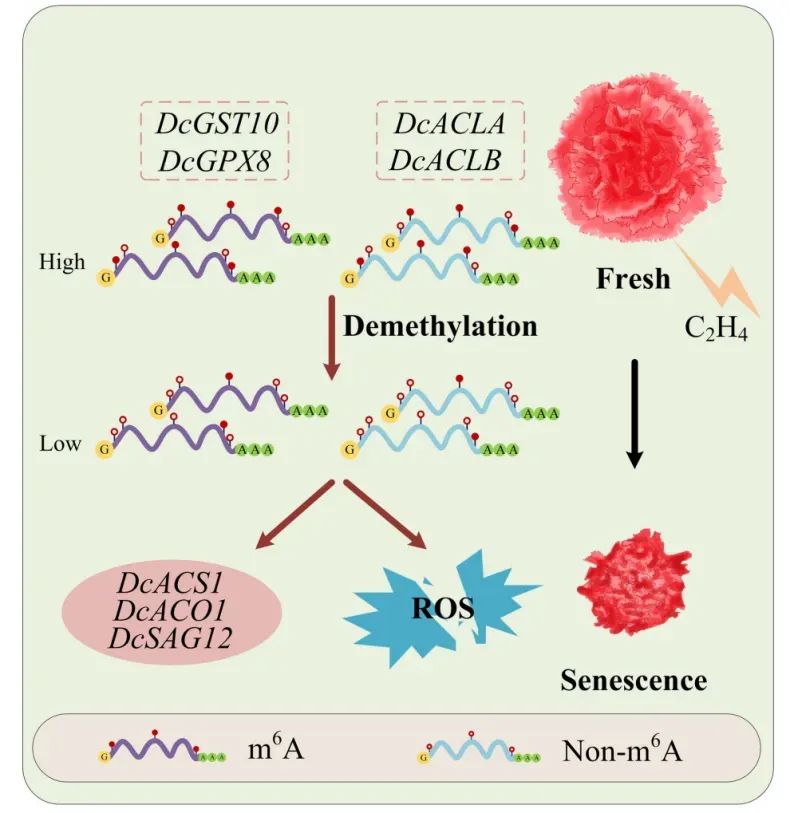
Fig9.m6A修饰调节康乃馨花衰老的模型图