酵母生产二氢槲皮素-文献精读117
Engineering Saccharomyces cerevisiae for the production of dihydroquercetin from naringenin
工程化酿酒酵母 (Saccharomyces cerevisiae) 以柚皮素 (naringenin) 生产二氢槲皮素 (dihydroquercetin)

摘要
背景 二氢槲皮素(DHQ)是一种强效的生物类黄酮,具有多种有益健康的特性,并在多种疾病的治疗中展现出潜力。二氢槲皮素的生物合成为满足其日益增长的需求提供了一种有前景的解决方案。然而,由于大量积累了橙皮苷(ERI)、柚皮素(NAR)、二氢山柰酚(DHK)及其他代谢产物,DHQ的生物合成产率较低,因此,这成为了DHQ生物合成的主要障碍。
研究结果 在本研究中,我们提出了几种策略以提高目标产物的生成并减少代谢物的积累。我们在酿酒酵母(S. cerevisiae)*中共表达了来自不同物种的*黄酮 3′-羟化酶(F3′H)*和*细胞色素 P450 还原酶(CPR),其中表达P450 还原酶酶复合体(SmF3′H/ScCPR)的最佳菌株在深孔微孔板实验中可将柚皮素(NAR)转化为435.7 ± 7.6 mg/L 的ERI。通过突变预测的泛素化位点以提高 SmF3′H 的稳定性,我们进一步提升了产物转化率,其中使用突变型 SmF3′H(K290R)后产量提高了12.8%。此外,我们测试了来自不同来源的黄酮 3-羟化酶(F3H)及其不同启动子对 DHQ 产量的影响,结果表明最佳菌株可将 1 g/L 的 NAR 转化为 381.2 ± 10.7 mg/L 的 DHQ,这表明对 F3H 表达进行时间调控对于最大化 F3′H 和 F3H 的功能至关重要。
结论 本研究提供了一种提高 NAR 向 DHQ 转化率的有效策略,并可作为相关研究的参考。
引言
二氢槲皮素(Dihydroquercetin,DHQ),又称紫杉酚(taxifolin),是一种具有生物活性的黄酮类化合物,广泛存在于水飞蓟、红洋葱、巴西莓等植物中,并可在 Legalon、Pycnogenol 和 Venoruton 等多种商业制剂中找到 [1, 2]。DHQ 具有两个立体中心,并存在四种对映异构体:( +)-taxifolin、(−)-taxifolin、( +)-epitaxifolin 和 (−)-epitaxifolin,它们在体内的活性和分布各不相同 [3]。DHQ 具有抗氧化、抗病毒和抗癌等多种生物活性,使其成为重要的膳食补充剂和功能性食品,并在神经退行性疾病、高血压和病毒感染等疾病的预防和治疗中展现出潜在的治疗价值 [2, 3]。研究表明,DHQ 的抗氧化能力优于生育酚和抗坏血酸等常见抗氧化剂,能够有效清除自由基,保护机体组织免受氧化损伤 [4]。在体内和体外研究中,DHQ 通过多种机制展现出抗癌效果,同时对正常细胞的副作用极小或几乎没有 [5]。近年来,研究表明 DHQ 通过抑制 3C 样蛋白酶(3CLpro)对冠状病毒具有潜在抑制作用 [6],并可能在未来作为有效的自由基清除剂用于 COVID-19 的治疗 [7]。随着人们对低毒高效药物需求的增长,DHQ 作为潜在的药物开发对象逐渐受到关注,以满足日益增长的市场需求。
目前,DHQ 主要从兴安落叶松(Dahurian Larch)*的木材中提取,但这种树木生长缓慢,主要分布在极端寒冷的北方地区 [8]。从植物材料中提取 DHQ 需要使用有机溶剂,过程繁琐且存在环境污染问题 [8,9,10]。化学合成 ( ±)-taxifolin 已有报道,但其光学拆分依赖于手性反相 HPLC,增加了成本和复杂性 [11]。虽然约 25% 的上市药物为外消旋混合物,但单一对映异构体通常具有更高的治疗价值,可能具有更好的生物利用度、更有效的治疗作用、更少的个体间差异,并能降低药物在代谢较弱个体中的累积 [12, 13]。相比化学合成,生物生产更适合合成此类具有立体选择性的复杂天然化合物,同时消费者对标有“生物”标签的产品更具偏好。DHQ 可通过*黄烷酮 3-羟化酶(F3H)*在 ERI 的 C 环 3′-位或 DHK 的 B 环 3-位进行立体特异性羟化来生成,而这两种前体均可由柚皮素(NAR)合成,而 NAR 可从果汁工业的废弃物中提取,成本较低 [14]。工程化*酵母 Yarrowia lipolytica 可利用葡萄糖生产 110.5 mg/L 的 DHQ,但同时积累了大量代谢物,包括 252.4 mg/L 的 NAR 和 134.2 mg/L 的 ERI [15]。最近的研究还显示,工程化大肠杆菌(Escherichia coli)可将 100 mg/L 的 NAR 转化为 13.6 mg/L 的 DHQ,进一步改造后可利用甘油生产 20.1 mg/L 的 DHQ,但仍存在大量 DHK 和 NAR 的积累 [16]。这些研究表明,DHQ 可通过 NAR 进一步羟化生成,但未来研究仍需解决代谢中间产物积累严重和目标产物产量低的问题。
已有研究表明,黄酮 3′-羟化酶(F3′H) 和 黄烷酮 3-双加氧酶(又称黄烷酮 3-羟化酶,F3H) 是 NAR 羟化的限速步骤 [15,16,17],其中 F3′H 依赖细胞色素 P450 还原酶(CPR)提供电子转移。目前,已鉴定并分析了多种 F3′H 和 F3H [18,19,20,21],但尚无系统比较其在 DHQ 生产中的活性研究。研究还表明,优化酶及其氧化还原伴侣的表达可显著提高目标产物的合成效率。例如,Lv 等研究发现查尔酮合酶(CHS)和细胞色素 P450 还原酶(CPR)是黄酮羟化的瓶颈,并通过调整 CHS 和 CPR 的基因拷贝数优化其表达,提高了黄酮类化合物的产量 [15]。另一项研究利用 CYP450 BM3 变体催化 NAR 向 ERI 的转化,在 3 L 生物反应器中,最佳突变体可将 100 μM NAR 转化为 47 μM ERI [22]。此外,Gao 等人通过调整启动子强度并突变 F3′H 及其氧化还原伴侣,成功在摇瓶实验中提高了 ERI 的产量,使其可由 1.5 g/L NAR 转化为 1 g/L ERI [23]。这些研究表明,提高 F3′H 和 F3H 的活性或优化其表达可有效促进黄酮类化合物的合成。
本研究通过优化 F3′H 与细胞色素 P450 还原酶(CPR) 的相互作用,提高底物转化效率,并通过修饰 F3′H 预测的潜在泛素化位点,提高其在细胞内的稳定性。此外,我们测试了不同来源的 F3H 及不同类型的启动子,以优化 DHQ 产量。在摇瓶实验中,最佳菌株可由 1 g/L 的 NAR 生产 381.2 ± 10.7 mg/L 的 DHQ。
材料与方法
菌株、质粒与引物
所有引物均由 GENEWIZ(中国) 和 Sangon Biotech(中国) 合成,具体序列列于 附加文件 1:表 S2。克隆实验采用 大肠杆菌 JM109(E. coli JM109),而用于基因表达和全细胞催化的酿酒酵母菌株 S. cerevisiae C800 由 S. cerevisiae CEN.PK2-1D(MATα, ura3-52, trp1-286, leu2-3, 112, his3Δ1)经 gal80 基因失活改造而得。本研究采用 pY26 质粒进行 染色体外表达。
培养基与培养条件
大肠杆菌菌株培养于锥形瓶中的 Luria–Bertani(LB) 培养基,在220 rpm、37 °C 振荡培养。质粒构建过程中,培养基需添加 100 μg/mL 氨苄青霉素(ampicillin) 以保持质粒稳定性(适用于液体培养基和固体培养基)。构建的质粒经 GENEWIZ(中国) 测序验证,并按照文献方法 [24] 转化至酵母细胞中。
对于重组酵母菌株的筛选,采用 酵母氮源培养基(YNB) 配制合成培养基,以分离营养缺陷型突变株,而 酵母提取物-蛋白胨-葡萄糖培养基(YPD) 用于酵母细胞培养。根据不同实验需求,酵母菌株可在 30 °C 下的 液体或固体 YNB 或 YPD 培养基 中培养。
表达质粒构建
基因片段经 GENEWIZ(中国) 进行密码子优化并合成,并采用 Gibson Assembly 方法,将其与 pY26-TEF-GPD [25] 和 pRS424 [26] 质粒骨架组装。构建的质粒经测序验证后,转化至酿酒酵母(S. cerevisiae) 用于生物转化。合成基因与构建的质粒 列于 附加文件 1:表 S1 和表 1。
| Constructed plasmids | Characteristics | Reference |
|---|---|---|
| pY26-SmF3′H-SmCPR | Episomal expression vector, containing the genes of SmF3′H and SmCPR from S. marianum | This study |
| pY26-SmF3′H-AtCPR | Episomal expression vector, containing the genes of SmF3′H from S. marianum and AtCPR from A. thaliana | This study |
| pY26-SmF3′H-EbCPR | Episomal expression vector, containing the genes of SmF3′H from S. marianum and EbCPR from E. breviscapus | This study |
| pY26-SmF3′H-GmCPR | Episomal expression vector, containing the genes of SmF3′H from S. marianum and GmCPR from G. max | This study |
| pY26-SmF3′H-HtCPR | Episomal expression vector, containing the genes of SmF3′H from S. marianum and HtCPR from H. tuberosus | This study |
| pY26-SmF3′H- HtCPR(L1M) | Episomal expression vector, containing the genes of SmF3′H from S. marianum and HtCPR from H. tuberosus(replaced the start codon UUG with ATG) | This study |
| pY26-SmF3′H- HtCPR(+ M) | Episomal expression vector, containing the genes of SmF3′H from S. marianum and HtCPR from H. tuberosus(added with a start codon of ATG) | This study |
| pY26-SmF3′H-ScCPR | Episomal expression vector, containing the genes of SmF3′H from S. marianum and HtCPR from S. cerevisiae | This study |
| pY26-FaF3′H-SmCPR | Episomal expression vector, containing the genes of FaF3′H from F. x ananassa and SmCPR from S. marianum | This study |
| pY26-FaF3′H-AtCPR | Episomal expression vector, containing the genes of FaF3′H from F. x ananassa and AtCPR from A. thaliana | This study |
| pY26-FaF3′H-EbCPR | Episomal expression vector, containing the genes of FaF3′H from F. x ananassa and EbCPR from E. breviscapus | This study |
| pY26-FaF3′H-GmCPR | Episomal expression vector, containing the genes of FaF3′H from F. x ananassa and GmCPR from G. max | This study |
| pY26-FaF3′H-HtCPR | Episomal expression vector, containing the genes of FaF3′H from F. x ananassa and HtCPR from H. tuberosus | This study |
| pY26-FaF3′H-ScCPR | Episomal expression vector, containing the genes of FaF3′H from F. x ananassa and ScCPR from S. cerevisiae | This study |
酵母基因组编辑
本研究采用酿酒酵母 CRISPR/Cas9 系统,按照文献方法 [27] 将 ScCPR 表达盒 整合至基因组 HO 位点。
具体方法如下:
-
通过 Benchling CRISPR gRNA Design Tool(CRISPR Guide RNA Design Tool | Benchling)分析并设计 sgRNA;
-
ScCPR 表达盒 与重组臂融合,并与 CRISPR/Cas9 质粒 共同转化至酵母细胞;
-
采用 YNB(不含尿嘧啶)合成培养基 筛选酵母突变株;
-
在 含 5-氟乳清酸(5-FOA) 的培养基中进行反向筛选,以移除 CRISPR/Cas9 质粒(对抗 URA3 基因)。
24 深孔板中的生物转化
-
重组菌株 先在 YNB-URA 培养基 中培养,挑取单菌落,接种至 10 mL YNB-URA 液体培养基,在 30 °C、220 rpm 培养至对数生长期;
-
向新鲜 YPD 培养基 中加入 250 μL 启动培养液,并补充 1 g/L NAR 作为底物,在 30 °C、220 rpm 条件下培养 72 h 后取样;
-
同样的方法,将 30 μL 启动培养液 接种至 3 mL 新鲜 YPD 培养基(含 1 g/L NAR),于 24 深孔微孔板 中培养,在 30 °C、220 rpm 培养 72 h 后取样分析。
分析方法
-
细胞生长情况 通过 微孔板读数仪(Tecan infinite 200 pro, Switzerland) 测定 光密度(OD 值),并进行适当稀释以确保测量范围合适;
-
代谢产物(NAR、DHQ、ERI、DHK) 提取:等体积纯甲醇混合样品,随后采用 岛津高效液相色谱(HPLC,Shimadzu Corporation, Kyoto, Japan) 进行定量分析;
-
HPLC 分析条件
:
-
色谱柱:Thermo Scientific Hypersil ODS-2 C18(Thermo Fisher Scientific Inc., USA);
-
流动相:含 0.1% 三氟乙酸(TFA) 的乙腈 和水,梯度洗脱;
-
流速:1.0 mL/min;
-
紫外检测波长(UV Detector A370):290 nm。
-
结果
黄酮 3′-羟化酶及其氧化还原伴侣的表达优化
黄酮类化合物 C 环和 B 环 在 3- 和 3′-位 的羟化反应分别由 黄烷酮 3-双加氧酶(F3H) 和 黄酮 3′-羟化酶(F3′H) 催化,该过程对于从 NAR(柚皮素,Naringenin) 生成 DHQ(双氢槲皮素,Dihydroquercetin) 至关重要(图 1A)。F3′H 属于细胞色素 P450 依赖型酶,能够将 NAR 羟化 生成 ERI(橙花素,Eriodictyol),但此过程需要氧化还原伴侣提供电子。
已有研究表明,通过优化 P450 酶与氧化还原伴侣的相互作用,可提高底物转化速率,增加产物得率 [28, 29]。这一优化可通过调整 F3′H 及其氧化还原伴侣的表达,促使其形成更具活性的酶复合体来实现。
本研究中,我们选择了以下来源的 CPR(细胞色素 P450 还原酶),并与 S. marianum(乳蓟,SmF3′H) 或 Fragaria × ananassa(草莓,FaF3′H) 的 F3′H 在酿酒酵母(S. cerevisiae) 中共表达(图 1B,表 1):
-
Silybum marianum(SmCPR,乳蓟)
-
Arabidopsis thaliana(AtCPR,拟南芥)
-
Erigeron breviscapus(EbCPR,灯盏细辛)
-
Glycine max(GmCPR,大豆)
-
Helianthus tuberosus(HtCPR,菊芋)
-
S. cerevisiae(ScCPR,酿酒酵母)
构建的重组菌株在 YPD 培养基(含 1 g/L NAR) 中培养 72 h 以进行酶表达,发酵结束后取上清液测定产物含量。
实验结果表明,SmF3′H 与 ScCPR、GmCPR 或 SmCPR 共表达的菌株 产生了更高滴度的 ERI。其中,SmF3′H/ScCPR 复合酶系统 的表现最佳,ERI 滴度达到 435.7 ± 7.6 mg/L,产率为 64.1 ± 2.2 mg/gCDW,在所有实验组中最高(图 1C)。
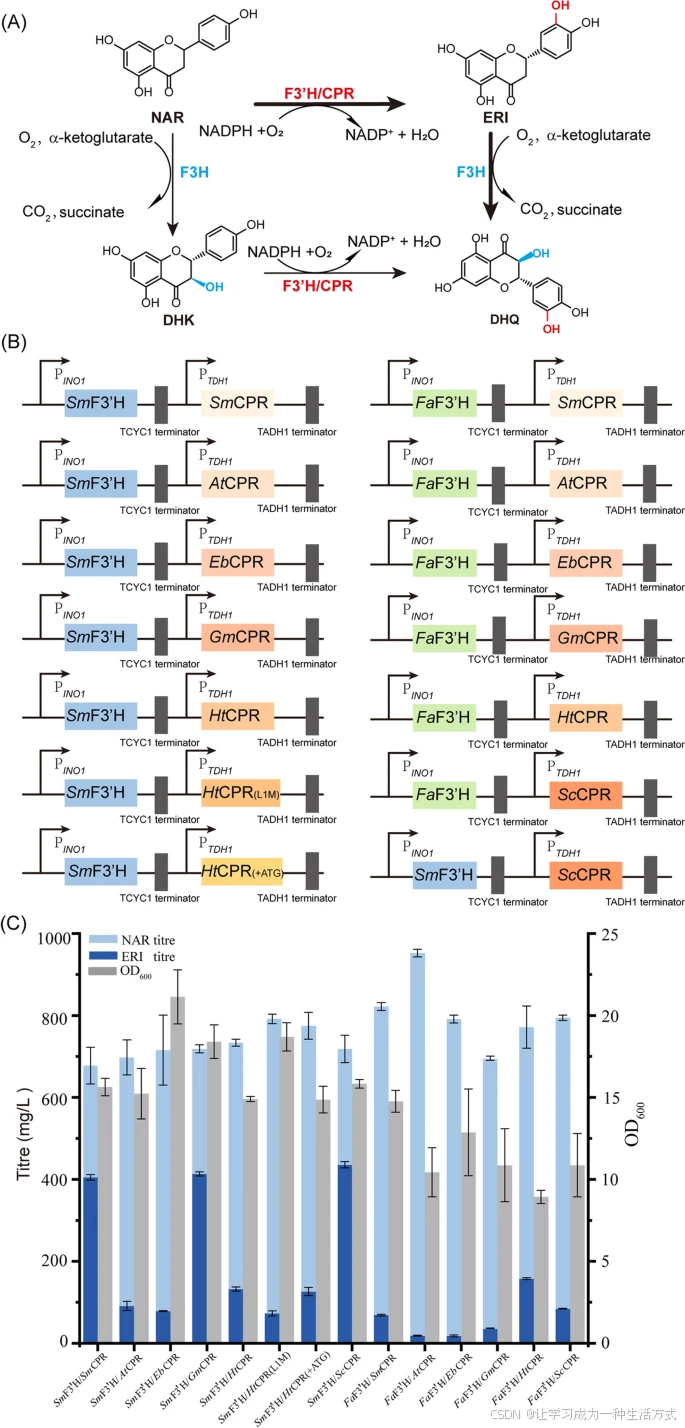
质粒构建及黄酮 3′-羟化酶与氧化还原伴侣的相互作用优化
A. 从 NAR 到 DHQ 的生物合成示意图 从 NAR 生成 DHQ 的生物合成示意图。
B. F3′H 与 CPR 共表达的质粒构建 用于 F3′H 和 CPR 共表达的质粒构建。
C. NAR 生物转化最优酶复合体的测试 所有来源不同的 F3′H 和 CPR 基因分别受 PINO1 和 PTDH1 启动子控制。 为了测试所表达的酶复合体活性,在 YPD 培养基中加入 1 g NAR 作为底物。 所有实验均进行三次重复,误差棒表示标准差(SD)。
截短作用于 SmF3′H 和 ScCPR,以研究跨膜区域的影响 利用 TMHMM-2.0 和 Alphafold 软件分析这些蛋白的跨膜区域,并预测其结构(附加文件 1:图 S1A)。 随后删除这些跨膜区域,得到不同的截短蛋白 SmF3′H-M1、SmF3′H-M2 和 ScCPR-M。 ScCPR 的跨膜区域删除后,对 NAR 到 ERI 的生物转化无显著影响。SmF3′H 的截短几乎完全消除了催化活性(附加文件 1:图 S1B)。
通过突变潜在泛素化位点提高酶的稳定性
在真核生物中,泛素介导的蛋白酶体降解是调节蛋白稳定性和更新的主要蛋白水解途径, 主要参与降解错误折叠、损伤的蛋白以及短寿命蛋白。 先前研究表明,对 FjTAL 中敏感的泛素化位点进行突变,可显著提高对羟基肉桂酸的产量。 为了鉴定泛素化位点并延长蛋白半衰期, 采用基于贝叶斯判别方法(Bayesian Discriminant method)的 BDM-PUB 在线服务器进行预测(表 2)。 将这些潜在泛素化位点突变为精氨酸,以构建 SmF3′H 突变体,并在酿酒酵母中以质粒方式表达, 该酵母菌株的 HO 位点已整合一份 ScCPR。重组菌株用于评估 SmF3′H 突变体在柚皮素到 ERI 转化中的贡献。 K290R 突变体能够略微提高 SmF3′H 在 NAR 到 ERI 转化中的表现,其产量相比野生型 SmF3′H 提高了 12.8%(图 2)。 其他预测泛素化位点的突变未能提高 ERI 产量,部分突变甚至导致产量下降, 表明这些突变在酶中产生了中性或有害影响。
| Protein | position | Sequence | scorces |
|---|---|---|---|
| SmF3′H | 244 | KMKKLHL | 2.13 |
| 290 | EGGKLSD | 1.58 | |
| 243 | KKMKKLH | 2.06 | |
| 92 | QFLKVHD | 1.90 | |
| 281 | ISLKDDA | 1.82 | |
| 108 | SGAKHIA | 1.23 | |
| 241 | VTKKMKK | 1.08 | |
| 427 | GGEKPNA | 1.05 | |
| 330 | QLLKQAQ | 3.04 | |
| 240 | SVTKKMK | 2.40 | |
| 482 | DPEKLNM | 0.51 | |
| 362 | AIVKETF | 2.15 |
-
The predicted ubiquitination sites were highlighted in bold
-
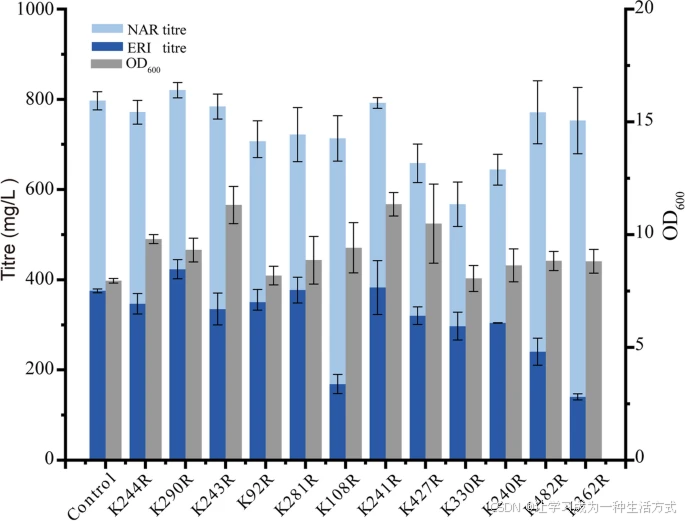
通过突变潜在的泛素化位点提高 SmF3′H 的稳定性 基于 BDM-PUB 预测的结果,对酶 SmF3′H 上的潜在泛素化位点进行了突变,并在整合了 ScCPR 基因至 HO 位点的 S. cerevisiae 菌株中进行瞬时表达。重组菌株在 72 小时发酵后,在 24 孔深孔板中测试其将 NAR 转化为 ERI 的能力。所有实验均进行三次重复,误差棒表示标准差(SD)。
优化黄烷酮 3-双加氧酶用于二氢槲皮素(DHQ)生产
酶 F3H 能够在 NAR 或 DHK 的 B 环 3 位羟基化这些化合物。从富含黄酮类化合物的不同植物物种中克隆了五种不同的 F3H,包括 S. marianum(SmF3H)、Citrus sinensis(CsF3H)、Epimedium koreanum Nakai(EkF3H)、G. max(GmF3H)和 Carthamus tinctorius(CtF3H)。为了测试它们的活性,这些 F3H 被插入含有 F3′H 和 ScCPR 的表达载体中,并在 S. cerevisiae 中受 PGAL7 启动子调控进行瞬时表达(图 3A)。
在 72 小时发酵后,表达 CsF3H 的重组菌株产生了 303.0 ± 1.7 mg/L 的 DHQ,产率为 66.0 ± 3.7 mg/gCDW,在所有测试组中表现最佳(图 3B)。表达 SmF3H 的重组菌株在 NAR 转化反应中也表现出较高活性,而表达 CtF3H 的菌株在 DHQ 生产方面表现最差。少量底物 NAR 在上清液中被检测到,但检测到了大量中间代谢物,包括 413.3 ± 7.8 mg/L 的 ERI 和 70.4 ± 7.2 mg/L 的 DHK(图 3B)。
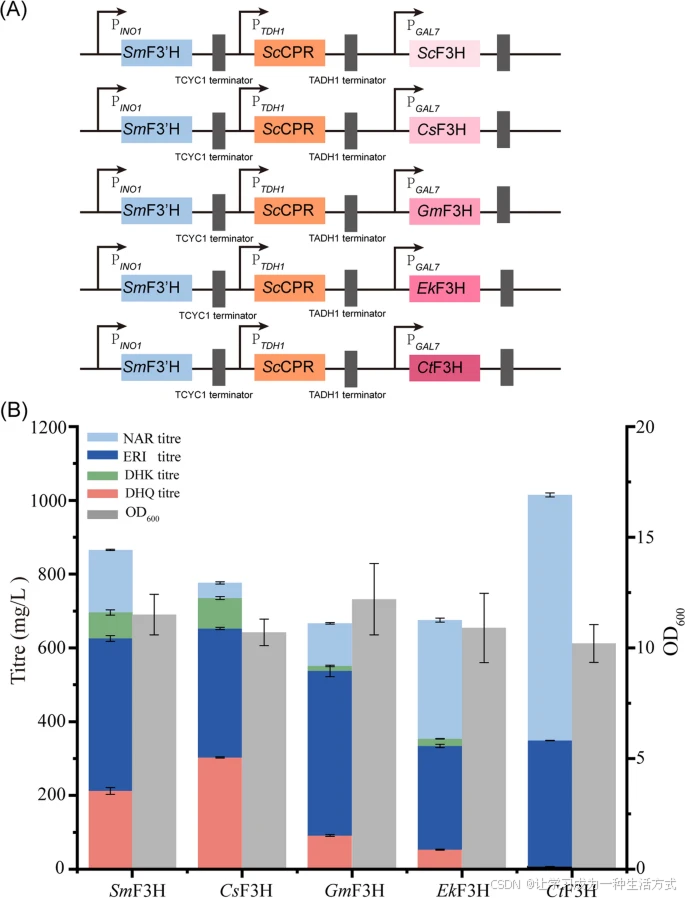
筛选最佳 F3H 用于二氢槲皮素(DHQ)生产 (A) 质粒构建用于共表达来自不同植物种类的 F3H 酶与 SmF3′H 和 ScCPR。F3H 的表达受可诱导启动子 PGAL7 调控。 (B) 通过在烧瓶中测定 NAR 的生物转化能力测试 F3H 的活性。菌株在补充 1 g/L NAR 作为底物的 YPD 培养基中培养。所有实验均进行三次重复,误差棒表示标准差(SD)。
通过启动子调控增强二氢槲皮素(DHQ)生产
上述研究表明,在 NAR 生物转化过程中积累了大量 ERI,表明 ERI 向 DHK 的转化效率不足。影响 F3H 表达和功能的因素可能包括 F3H 和 F3′H 之间因细胞内资源竞争导致的相互干扰、启动子强度的差异以及环境压力和 pH 变化引起的酶表达波动等 [33]。
基于这些考虑,研究采用了不同强度的组成型或可诱导启动子(如 PSED1、PTDH3、PCCW12、PGAL1、PGAL2、PGAL10 和 PHXT7)来调控 S. cerevisiae 中 F3H 的表达,并利用这些启动子构建了 F3H 的瞬时表达载体(图 4A)。按照先前的发酵方案进行实验,结果表明在所有测试组中,PGAL2 启动子的表现最佳。
采用 PGAL2 启动子调控 F3H 表达的重组菌株,从 1 g/L NAR 中积累了 381.2 ± 10.7 mg/L DHQ,产率为 67.0 ± 6.7 mg/gCDW,转化率达到 0.34 ± 0.005 molDHQ/molNAR,相比使用 Gal7 启动子的亲本菌株提高了 27.1%(图 4B)。
令人意外的是,强启动子 PSED1、PTDH3 和 PCCW12 在本研究中表现较差,并检测到大量底物 NAR,表明 F3′H 催化受到了干扰。另外,使用 PHXT7 启动子(一个在低浓度葡萄糖下诱导的葡萄糖依赖型启动子)导致了 ERI 的大量积累,而 DHQ 产量较低(图 4B)。

通过启动子调控增强二氢槲皮素(DHQ)生产 原始 PGAL7 启动子被不同的启动子替换,包括组成型启动子 PSED1、PTDH3、PCCW12 和 PHXT7,以及可诱导启动子 PGAL1、PGAL2 和 PGAL10。菌株在 YPD 培养基中与 SmF3′H、ScCPR 和 CsF3H 共同表达,并补充 1 g/L NAR 进行 DHQ 生产测试。所有实验均进行三次重复,误差棒表示标准差(SD)。
讨论
二氢槲皮素(DHQ)是一种具有生物活性的黄酮类化合物,因其多种健康益处而受到工业界和代谢工程领域的广泛关注。DHQ 的生物合成是一种有吸引力的替代方案,以满足日益增长的市场需求。目前,DHQ 可通过葡萄糖或甘油的 de novo 合成方式生产。在生物反应器中,S. cerevisiae 可生产约 330 mg/L DHQ [17],而在摇瓶水平,Y. lipolytica 仅能生产约 110 mg/L [15]。其产量较低的部分原因是代谢中间体的过量积累以及 de novo 合成途径较长导致的低效率,这需要深度代谢工程手段来平衡模块并有效引导 DHQ 合成的代谢流,同时降低代谢负担。此外,高附加值的 DHQ 还可利用低成本的 NAR(从果皮废料中提取)进行生产。本研究采用了多种策略来提高 NAR 向 DHQ 的转化率,包括优化 F3'H 与其氧化还原伴侣的相互作用、提高 F3'H 酶的稳定性、筛选最佳 F3H 以及对关键酶进行时序调控。这些改造的重组细胞不仅可作为 de novo DHQ 生物合成的可行方案,还可用于将低成本的 NAR 转化为高附加值产品 DHQ。
DHQ 的生物合成涉及黄酮类化合物 B 环在 3′-位的羟基化,该反应由细胞色素 P450 依赖型黄酮 3′-羟化酶(F3'H)催化,并需要细胞色素 P450 还原酶(CPR)提供电子。P450 依赖型 F3'H 和 CPR 形成 1:1 的功能性 P450-还原酶复合物,但 P450 酶和还原酶的平衡可能并不理想 [34]。此外,F3'H 和 ScCPR 作为膜结合蛋白,优化其表达水平有助于更有效地利用有限的膜空间及其他生物资源以促进蛋白质合成。先前的研究表明,通过调整 F3'H 和 CPR 的基因拷贝数或启动子强度来改变其相对表达水平,可有效提高 DHQ 产量 [15, 23]。当 P450 酶与还原酶的亲和力较高时,电子将优先流向这些 P450 [35]。因此,增强 F3'H 与其还原酶伴侣的相互作用有助于加速该过程。本研究表明,来自 S. marianum(SmF3'H)的 F3'H 和来自 S. cerevisiae(ScCPR)的 CPR 可形成功能性 P450-还原酶嵌合复合体,在 NAR 转化实验中表现最佳。此外,进一步的截短研究表明,SmF3'H 的跨膜区域对其功能维持至关重要,这可能是因为催化反应需要膜作为相互作用和反应的基质,而酶的组织和功能受膜成分调控 [36, 37]。
改变潜在泛素化位点也是提高 DHQ 产量的另一种策略。该方法可延长关键酶的半衰期,从而提高蛋白质稳定性并增加目标产物的合成。此前,我们的研究表明,通过改变 FjTAL 的泛素化敏感位点,可显著提高对羟基肉桂酸的合成 [32]。本研究对 SmF3'H 进行了类似的改造,突变泛素化敏感位点后,DHQ 产量略有提升。然而,一些突变导致了酶活性下降,可能是因为突变发生在催化活性区域或影响了其与氧化还原伴侣的相互作用。此外,细胞会通过泛素-蛋白酶体途径监测并降解错误折叠或异常蛋白 [38, 39],而错误折叠的 SmF3'H 可能形成聚集体,未能及时降解,从而对细胞产生一定干扰。值得庆幸的是,本研究中未观察到明显的细胞生长受损现象。
此外,我们还尝试提高 F3H 的活性和表达水平,F3H 是负责催化黄酮 C 环 3-位羟基化的关键酶。本研究筛选了 5 种来源于不同植物的 F3H,其中 Petroselinum crispum 和 A. thaliana 的 F3H 对 NAR 具有较高亲和力,但在温和 pH 条件下活性较低,因此未被选用 [40, 41]。本研究使用不同强度的启动子评估其对 DHQ 产量的影响。值得注意的是,使用强启动子 PSED1、PTDH3 和 PCCW12 [42] 反而导致 DHQ 产量显著下降,而与半乳糖代谢相关的启动子(如 PGAL 系列)表现优于上述三者。这表明 SmF3'H/CPR 的表达和/或功能可能受到了干扰。此外,不同来源的 F3'H 和 F3H 可能在催化不同底物时表现出不同的效率,并对某些底物具有偏好性 [43]。研究发现,SmF3'H 更倾向于使用 NAR 作为底物,而非 DHK,这意味着 DHQ 主要是通过 NAR 先转化为 ERI,再转化为 DHQ。在这种情况下,强组成型表达 F3H 可能会与 SmF3'H/CPR 表达竞争有限的细胞资源,并加重细胞合成酶的负担。而使用较弱的组成型启动子 PHXT7 促进了 NAR 向 ERI 的转化,但 DHQ 产量较低。为了避免这种可能的干扰,我们采用 PGAL1、PGAL2、PGAL10 和 PGAL7 这些可诱导启动子来调控 F3H 的表达。结合上述策略,DHQ 转化率较以往研究有了显著提升。
已有研究报道采用不同的菌株工程策略和发酵优化方法,使 S. cerevisiae 在生物反应器中通过 108 小时的补料分批发酵,DHQ 产量达到 330 mg/L,是烧瓶水平的三倍,也是目前报道的最高产量 [17]。另一项研究中,Y. lipolytica 工程菌在摇瓶水平生产 DHQ 110 mg/L,同时积累了较多的代谢中间体 [15]。本研究通过多种优化策略,在深孔微孔板中从 NAR 生产了 381.2 mg/L DHQ。值得注意的是,在生物反应器中,菌体密度远高于深孔微孔板,可能提高 9-10 倍。此外,进一步优化发酵条件仍可能提升 DHQ 产量。因此,本研究所开发的 NAR 生物转化菌株在 DHQ 生产方面具有更高的效率。然而,未来仍需在受控生物反应器中进一步验证和比较其性能。
结论
综上所述,本研究通过代谢工程改造 S. cerevisiae,使其能够将柚皮素(NAR)转化为二氢槲皮素(DHQ)。在 S. cerevisiae 中共表达了来自不同物种的关键酶 F3′H 及其 P450 还原酶伴侣,其中 S. marianum 的 F3′H(SmF3′H)与 S. cerevisiae 的 CPR(ScCPR)形成了最佳的 P450-还原酶复合体,可将 NAR 转化为 435.7 ± 7.6 mg/L 的 ERI,转化率为 0.41 ± 0.007 mol ERI/mol NAR。此外,通过突变预测的潜在泛素化位点,提高了 SmF3′H 的稳定性,进一步提升了产物生成效率。为了提高 DHQ 产量,本研究筛选了来自不同植物的 5 种 F3H,并与 SmF3′H/ScCPR 共同表达,同时优化了启动子组合,使 1 g/L NAR 的转化可产生 381.2 ± 10.7 mg/L DHQ。与先前研究相比,结合所有这些策略后,DHQ 的产量和转化率均得到显著提升。本研究提供了一套系统性策略,以优化 NAR 转化 DHQ 的生物合成过程。
缩写
-
DHQ: 二氢槲皮素(Dihydroquercetin)
-
ERI: 俄瑞克酮(Eriodictyol)
-
DHK: 二氢杨梅素(Dihydrokaempferol)
-
NAR: 柚皮素(Naringenin)
-
F3′H: 黄酮 3′-羟化酶(Flavonoid 3′-hydroxylase)
-
F3H: 黄烷酮 3-羟化酶(Flavanone 3-hydroxylase)
-
CHS: 查尔酮合酶(Chalcone synthase)
-
CPR: 细胞色素 P450 还原酶(Cytochrome P450 reductase)
-
CRISPR: 成簇规律间隔短回文重复序列(Clustered Regularly Interspaced Short Palindromic Repeat)