金丝桃属植物中呫吨酮的生物合成提供苯甲酰辅酶 A--文献精读173
A promiscuous coenzyme A ligase provides benzoyl-coenzyme A for xanthone biosynthesis in Hypericum
一种非特异性辅酶 A 连接酶为金丝桃属植物中呫吨酮的生物合成提供苯甲酰辅酶 A

摘要
苯甲酸衍生物(如多异戊烯基二苯甲酮与呫吨酮)因其复杂的化学结构与多样的生物活性,受到科研人员的广泛关注。金丝桃属(Hypericum)植物具有极高的药用价值,贯叶金丝桃(H. perforatum)便是典型代表。该属植物富含二苯甲酮与呫吨酮衍生物,其生物合成需苯甲酸 - 辅酶 A(benzoate-CoA)连接酶(BZL)催化 —— 该酶可将苯甲酸活化生成苯甲酰辅酶 A(benzoyl-CoA)。
尽管目前对植物中苯甲酸生物合成的研究已取得显著进展,但此前所有关于 BZL 基因与蛋白的结构研究,均局限于降解苯甲酸的微生物。本研究利用转录组资源,从产呫吨酮的蜡菊(Hypericum calycinum)细胞培养物中,克隆得到一个植物酰基激活酶(AAE)的转录本,命名为HcAAE1。
诱导子处理后,HcAAE1的转录水平先于呫吨酮积累而升高,这一规律与该途径中其他相关基因的表达模式一致。报告基因融合蛋白的亚细胞定位结果显示,由于存在 2 型过氧化物酶体靶向信号,HcAAE1可同时定位于细胞质与过氧化物酶体。这一结果表明,金丝桃属植物中苯甲酰辅酶 A 的生成,依赖于辅酶 A(CoA)依赖性的非 β- 氧化途径。
基于荧光素酶的底物特异性检测与动力学分析表明,HcAAE1具有非特异性底物偏好,其中苯甲酸是其唯一可接受的芳香族底物。与 4 - 香豆酸 - CoA 连接酶和肉桂酸 - CoA 连接酶不同,HcAAE1无法分别接受 4 - 香豆酸与肉桂酸作为底物。
计算机模拟建模进一步验证了这一底物偏好:模型显示,苯甲酸及其单磷酸腺苷中间体均可在HcAAE1(即 BZL)的活性位点腔中有效对接。
引言
苯甲酸是植物来源的多种特化代谢产物的关键生物合成前体(图 1)。在产该物质的植物体内,这些成分主要发挥化学防御功能,例如苹果中的奥古巴宁(aucuparin)与拟南芥中的 3 - 苯甲酰氧基丙基硫代葡萄糖苷(Kliebenstein 等,2007;Beerhues 与 Liu,2009)。
此外,苯甲酸衍生物还包括多种具有重要药理活性的物质:例如临床应用广泛的抗癌化合物紫杉醇(商品名 Taxol;Barbuti 与 Chen,2015),以及麻醉药物可卡因(Colzato 等,2009)。苯甲酸衍生物中还包含呫吨酮类化合物,这类物质具有强效抗菌与抗真菌活性(Tocci 等,2013;Simonetti 等,2016;Badiali 等,2018;Gaid 等,2020)。
在金丝桃属植物中,呫吨酮主要在根部积累,为植物抵御土传病原体提供化学屏障(Tocci 等,2018)。其生物合成机制多年来一直是研究热点(Gaid 等,2012;Fiesel 等,2015;El-Awaad 等,2016;Nagia 等,2019;Khattab 与 Farag,2020)。由于蜡菊(H. calycinum)细胞悬浮培养物与贯叶金丝桃(H. perforatum)生长素诱导根培养物的呫吨酮积累量高于田间种植植株,二者被用作研究呫吨酮核心结构生物合成的模式系统(El-Awaad 等,2016;Tocci 等,2018)。
未处理的蜡菊细胞培养物不含呫吨酮;但经酵母提取物处理后,细胞中总呫吨酮含量可达约 4 mg/g 干生物量(Gaid 等,2012)。近年来,研究人员已从经诱导子处理的蜡菊细胞培养物中,克隆得到多个与异戊烯基呫吨酮生物合成相关的下游编码序列(Fiesel 等,2015;El-Awaad 等,2016;Nagia 等,2019)。
本研究利用该实验系统,探究呫吨酮骨架组装的起始底物 —— 苯甲酰辅酶 A(benzoyl-CoA)的生物合成机制(呫吨酮骨架组装需苯甲酰辅酶 A 与丙二酰辅酶 A 共同参与)。尽管苯甲酸结构简单,但其衍生的苯甲酰辅酶 A 可通过多种并行途径生成,这使得其生物合成机制的完整解析面临挑战(Wildermuth,2006;Widhalm 与 Dudareva,2015)。
然而,过去二十年间的大量研究已显著提升了人们对植物中苯甲酰辅酶 A 生物合成的认知(Abd El-Mawla 与 Beerhues,2002;Gaid 等,2012;Saini 等,2017;Park 等,2017;Saini 等,2019;Teotia 等,2019;Saini 等,2020)。在矮牵牛(Petunia hybrida)中,这一领域的研究尤为深入(Boatright 等,2004;Orlova 等,2006;Van Moerkercke 等,2009;Colquhoun 等,2012;Klempien 等,2012;Qualley 等,2012;Adebesin 等,2018)。
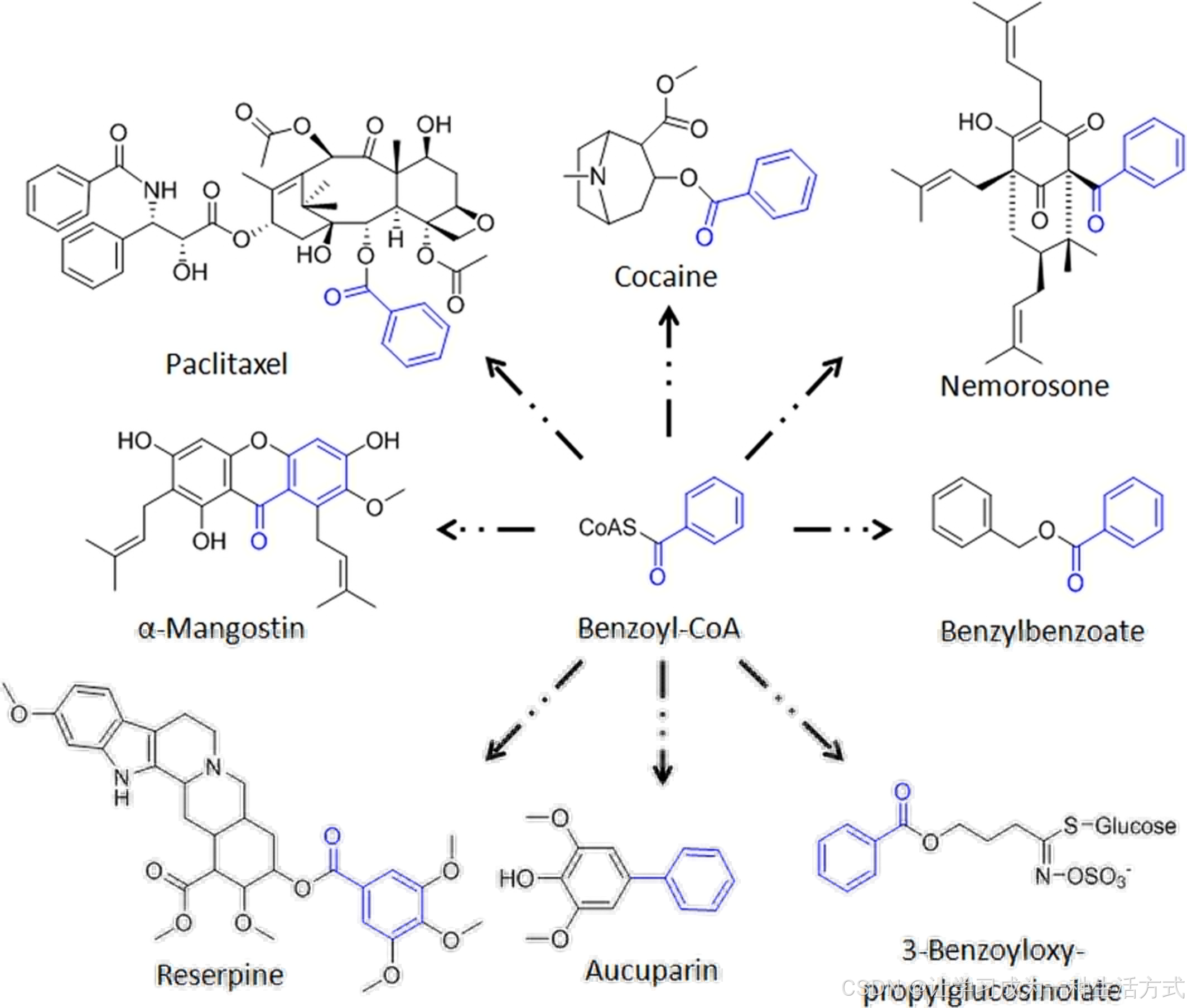
图 1 具有生物活性的苯甲酰基衍生天然产物示例
紫杉醇(Paclitaxel)用于治疗多种癌症(Barbuti 与 Chen,2015);可卡因(cocaine)可用作局部麻醉剂,也是一种成瘾性毒品(Colzato 等,2009);海滨霉素(nemorosone)对肿瘤细胞具有潜在细胞毒性(Simpkins 等,2012);α- 倒捻子素(α-mangostin)是抑制癌症发生的化学预防剂(Zhang 等,2017);利血平(reserpine)长期用于治疗高血压(Shamon 与 Perez,2016);苯甲酸苄酯(benzylbenzoate)是治疗人类疥疮的外用药物(Bachewar 等,2009);奥古巴宁(aucuparin)与 3 - 苯甲酰氧基丙基硫代葡萄糖苷分别是苹果与拟南芥的防御化合物(Kliebenstein 等,2007;Beerhues 与 Liu,2009)。
苯甲酸的生物合成主要通过两条途径:一是莽草酸 / 分支酸途径;二是核心苯丙烷途径 —— 该途径通过缩短肉桂酸侧链的 2 个碳原子(C2 缩短)生成苯甲酸(Wildermuth,2006;Widhalm 与 Dudareva,2015)。核心苯丙烷途径又可进一步分为三类:辅酶 A(CoA)依赖性 β- 氧化途径、CoA 依赖性非 β- 氧化途径,以及 CoA 非依赖性非 β- 氧化途径。
这三类途径的起始步骤相同:均由苯丙氨酸解氨酶(PAL)催化 L - 苯丙氨酸脱氨,生成反式肉桂酸;反式肉桂酸的活性形式为肉桂酰 - CoA,该物质在 β- 氧化途径中于过氧化物酶体内被氧化,在非 β- 氧化途径中则于过氧化物酶体外被氧化。此外,游离反式肉桂酸的氧化也可独立于过氧化物酶体 β- 氧化途径进行。
CoA 非依赖性非 β- 氧化途径的关键步骤是:通过直接缩短反式肉桂酸的 C2 侧链生成苯甲醛,苯甲醛随后被氧化为苯甲酸(Saini 等,2019)。植物中苯甲酰 - CoA 的生成可同时存在一条或多条交叉途径(Boatright 等,2004;Van Moerkercke 等,2009;Widhalm 与 Dudareva,2015)。
在腺柱金丝桃(Hypericum androsaemum)细胞培养物中,已从生化水平检测到 CoA 依赖性非 β- 氧化途径的存在(图 2;Abd El-Mawla 与 Beerhues,2002)。该途径的起始步骤为:肉桂酸 - CoA 连接酶(CNL)催化反式肉桂酸转化为肉桂酰 - CoA。2012 年,研究人员从蜡菊(H. calycinum)、拟南芥(Arabidopsis thaliana)与矮牵牛(P. hybrida)中克隆并鉴定出 CNL 的 cDNA,发现其编码的蛋白定位于过氧化物酶体(Gaid 等,2012;Klempien 等,2012;Lee 等,2012)。
腺柱金丝桃的无细胞提取物可将肉桂酰 - CoA 的侧链缩短 2 个碳原子:先形成 3 - 羟基 - 3 - 苯基丙酰 - CoA 中间体,再生成苯甲醛。该转化过程被认为由肉桂酰 - CoA 水合酶 / 裂解酶的双功能活性催化(Abd El-Mawla 与 Beerhues,2002),但目前尚未明确编码该酶的 cDNA 及其蛋白的亚细胞定位。
此前,研究人员从革兰氏阴性菌荧光假单胞菌(Pseudomonas fluorescens)中纯化出 4 - 羟基肉桂酰 - CoA 水合酶 / 裂解酶,该酶可催化 4 - 羟基肉桂酰 - CoA 底物的水合反应与逆醛缩合裂解反应,但无法作用于肉桂酰 - CoA(Mitra 等,1999)。之后,拟南芥中Chy1基因的 cDNA 被克隆,该基因编码的蛋白是 4 - 羟基肉桂酰 - CoA 水合酶 / 裂解酶的远缘同源物;但体外实验显示,该酶仅对肉桂酰 - CoA 具有水解活性,无法生成苯甲醛(Ibdah 与 Pichersky,2009)。
CoA 依赖性非 β- 氧化途径的倒数第二步,是由 NAD + 依赖性酶 —— 苯甲醛脱氢酶(BD)催化苯甲醛转化为苯甲酸。研究人员已在腺柱金丝桃、花楸(Sorbus aucuparia)与梨(Pyrus pyrifolia)的细胞培养物无细胞提取物中检测到 BD 活性,其中花楸与梨可合成奥古巴宁(Abd El-Mawla 与 Beerhues,2002;Gaid 等,2009;Saini 等,2017);此外,还对金鱼草(Antirrhinum majus)中定位于线粒体的重组 BD 蛋白进行了鉴定(Long 等,2009)。
金丝桃属植物中该途径的最后一步,是由苯甲酸 - CoA 连接酶(BZL)催化苯甲酸活化生成苯甲酰 - CoA,这一步骤此前仅在生化水平得到证实(Abd El-Mawla 与 Beerhues,2002)。而在矮牵牛中,CoA 依赖性 β- 氧化途径的辅助酶 —— 过氧化物酶体硫酯酶,可将该途径在过氧化物酶体内生成的苯甲酰 - CoA 转化为苯甲酸(图 2;Qualley 等,2012;Adebesin 等,2018)。
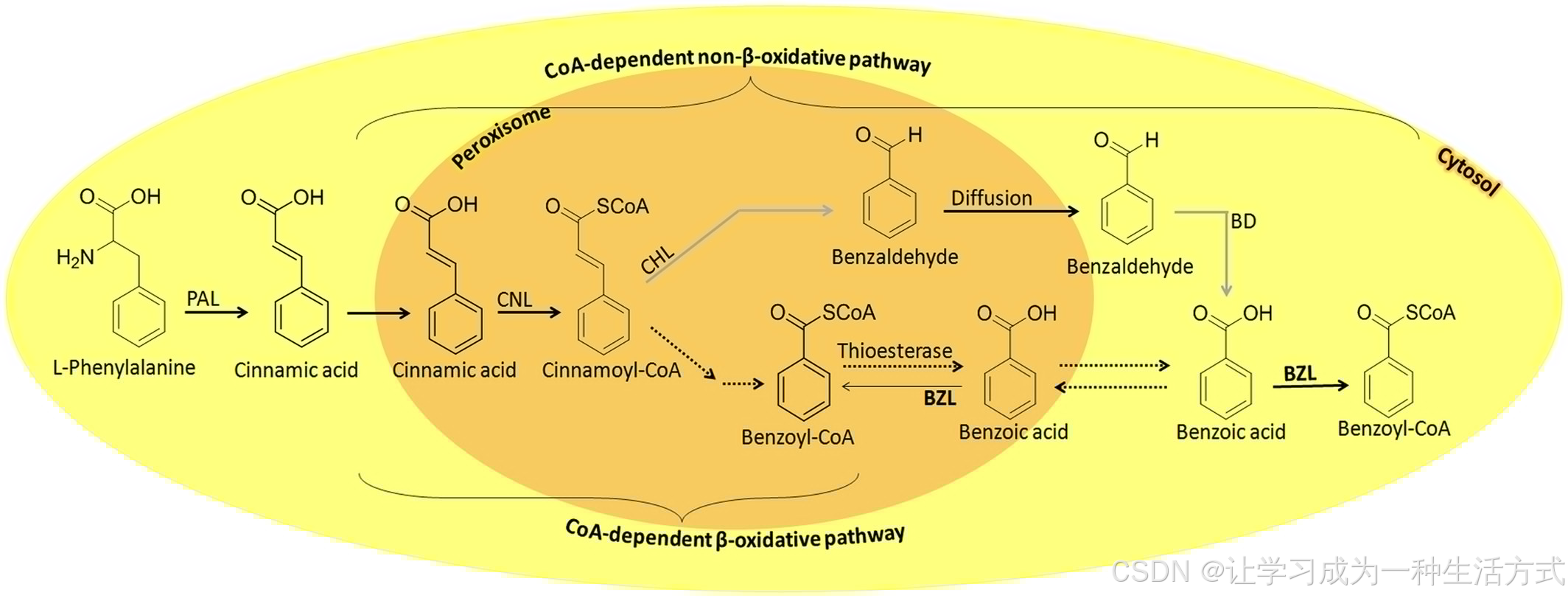
图 2 金丝桃属植物中苯甲酰辅酶 A(benzoyl-CoA)的推测生物合成途径 —— 该物质可存在于细胞质中供下游反应利用
苯丙氨酸解氨酶(PAL)催化反应后,两条途径被框出:一是辅酶 A(CoA)依赖性非 β- 氧化途径;二是 “CoA 依赖性 β- 氧化途径 + 裂解、转运与再活化” 途径,这两条途径均依赖两种辅酶 A 连接酶(肉桂酸 - CoA 连接酶 CNL 与苯甲酸 - CoA 连接酶 BZL)。图中标注了各反应的亚细胞区室定位及中间产物的转运方向。(缩写说明:PAL,苯丙氨酸解氨酶;CNL,肉桂酸 - CoA 连接酶;CHL,肉桂酰 - CoA 水合酶 / 裂解酶;BD,苯甲醛脱氢酶;BZL,苯甲酸 - CoA 连接酶。)灰色箭头代表已在酶水平证实但未明确基因的步骤;虚线箭头代表推测步骤。
苯甲酸 - CoA 连接酶(BZL)属于腺苷酸形成酶超家族中的酰基激活酶(AAE)(Fulda 等,1994;Schmelz 与 Naismith,2009),可通过 ATP 依赖性的两步反应激活苯甲酸(图 S1a)。目前,仅微生物来源的 BZL 在基因与蛋白结构水平得到深入研究(Egland 等,1995;Xiang 与 Moore,2003;Thornburg 等,2015);而被子植物的 BZL 仅在生化水平被研究(Abd El-Mawla 与 Beerhues,2002;Beuerle 与 Pichersky,2002;Saini 等,2020)。
最近有研究发现,裸子植物云南红豆杉(Taxus media)的一种酰基激活酶(TmAAE4)对底物具有特异性,仅能作用于苯甲酸,推测其参与紫杉醇侧链的生物合成(Srividya 等,2020)。尽管针对苯甲酸的并行及交叉生物合成途径已有大量研究,但在基因与结构水平上支持植物中苯甲酸酶促激活过程的证据仍十分稀缺且不完整。
在金丝桃属植物中,BZL 是推动苯甲酸流向呫吨酮生物合成途径的关键酶。此前研究已利用产呫吨酮的蜡菊(H. calycinum)细胞培养物,分离得到与 CoA 依赖性非 β- 氧化途径相关的 CNL 转录本(Gaid 等,2012)。因此,该模式系统也适用于分离 BZL 转录本。本研究报道了蜡菊酰基激活酶HcAAE1的 cDNA 克隆、功能与动力学鉴定、表达分析、亚细胞定位及同源建模结果,证实该酶可高效激活苯甲酸。
结果
克隆与生物信息学分析获得两个HcAAE cDNA
以蜡菊肉桂酸 - CoA 连接酶 HcCNL(登录号 AFS60176.1)为查询序列,从公开的贯叶金丝桃(H. perforatum)转录组数据库 ——“千种植物转录组计划” 数据库(OneKP:https://sites.google.com/a/ualberta.ca/onekp)中检索假定的 BZL 转录本。
设定筛选标准如下:检索与 HcCNL 匹配度显著且相似度为 50%-55% 的重叠群(contig),同时排除与 4 - 香豆酸 - CoA 连接酶(4CL)及长链酰基 - CoA 合成酶(LACS)同源性≥40% 的序列(Shockey 等,2002)。该筛选依据源于此前的系统发育分析结果:假定的 BZL 与 CNL 同属 VI 进化枝,而 4CL 与 LACS 分别归属于 IV 进化枝与 I 进化枝(Shockey 与 Browse,2011;Gaid 等,2012;Klempien 等,2012)。
通过 “每百万映射片段中每千碱基长度的片段数”(FPKM)值,进一步筛选检索到的序列(MPGR 方法)—— 金丝桃属植物中呫吨酮在根部积累,因此根部高表达的序列被优先选择。应用上述标准,最终筛选得到 5 条序列(表 S2),均包含腺苷酸形成酶超家族特有的 10 个核心基序(Gulick,2009),包括 BOX I 基序、ATP / 单磷酸腺苷(AMP)结合基序(序列为 SSGTTXXPKGV,PROSITE 数据库登录号 PS00455)及 BOX II 基序(序列为 GEIXXXG)(Ehlting 等,2001;图 S2)。
其中两条重叠群(BNDE_scaffolds 2018166 与 2018167)对应同一 cDNA(表 S2)。以经诱导子处理的蜡菊细胞培养物的逆转录 RNA 为模板,利用基因特异性引物(GSP)(表 S1),克隆得到HcAAE1与HcAAE2的开放阅读框(ORF)。HcAAE1与HcAAE2的 ORF 长度分别为 1647 bp 与 1692 bp。
采用标准 PCR 与降落 PCR(touch-down PCR),均未检测到数据库中另外两条序列(onekp:BNDE_scaffold_2026015,对应引物 GSP3+4;onekp:BNDE_scaffold_2009069,对应引物 GSP5+6)的转录本(表 S1、S2)。
HcAAE1与HcAAE2的 OR