生物化学Learning Track(17)脂质
本笔记基于杨荣武教授主编的《生物化学》第四版,配图也来自于教材
1. 脂质的结构和功能
脂质根据化学结构可以分为三大类,即简单脂、复合脂和异戊二烯类脂
一、简单脂
简单脂包括游离的脂肪酸(FFA)以及由FFA和醇形成的酯(包括脂肪和蜡)。其中脂肪是甘油三酯(TG)
按照碳原子数目,FFA可以分为奇数脂肪酸和偶数脂肪酸,但是天然的FFA大多是偶数脂肪酸(因为自然界当中的脂肪酸是以两个单位的烃基相加的)。FFA根据碳氢链的饱和度又可以分为饱和脂肪酸和不饱和脂肪酸。天然不饱和脂肪酸的双键多为顺式(有利于维持膜的流动性,使得脂肪酸链之间难以靠近),某些细菌含有反式的不饱和脂肪酸。根据对于动物的营养价值,脂肪酸还分为必需脂肪酸和非必需脂肪酸(定义和氨基酸是一致的)。其中亚油酸和α-亚麻酸是必需脂肪酸(亚油酸是合成花生四烯酸的前体,花生四烯酸又是多种前列激素合成的前体,α-亚麻酸是DHA和EPA的前体)
脂肪最重要的生理功能是贮存能量,其贮能的效率要高于糖原和蛋白质(因为脂肪的还原程度更高,消耗更多的氧从而获得更多的能量)
脂肪作为能量储备的时候,一般贮存在白色脂肪细胞当中,与胆固醇酯或者其他固醇酯一起以脂滴的形式存在。在脂滴的表面是一层磷脂单分子层(因为内部是脂溶性分子,如果是水溶性分子那么就是双分子层形成的脂质体),在外面还包被一层脂外被蛋白(PLIN)PL你广泛存在于各种动物的脂肪细胞当中,在脂滴的表面形成了一层保护性的屏障,可以防止脂肪酶对脂肪的水解。
但是如果收到外部信号的激活(比如肾上腺素和胰高血糖素),那么PKA就会被激活,PKA会修饰PLN使其失去屏障作用,同时激活脂肪酶,从而开始水解脂肪。
我们在分析糖原为什么适合作为储能物质的时候说到两点,一点是这种多聚物储存能量对于渗透压的影响很小,另外一点是糖原具有高度的分支结构,这样就使得糖原具有很多的非还原端可以被用来作为水解酶的结合位点,即可开始水解。
那么对于脂肪而言,除了其储存在脂滴当中不会对渗透压产生影响以及具备很高的能量密度之外,它不易被动员水解,对于糖原的水解直接激活酶即可(糖原是水溶性的),但是对于脂滴需要发生屏障的打开和脂肪酶的激活,同时分布也并不广泛,但是这一点也恰好适配它作为长期储能物质的生物功能。但是能量密度高的前提是可以释放出能量,虽然说脂肪的还原度高,但是这也就意味着需要更多的氧气才可以将脂肪分解放能,所以脂肪在无氧和缺氧的情况下也是发挥不了作用的。(另外,由于脂肪的H含量很高,所以其氧化的过程也会产生很多代谢水,实际上骆驼就是采用储存脂肪的方式来储存水资源)
(反式脂肪酸的定义是不饱和脂肪酸的双键是反式的,自然界当中正常的不饱和脂肪酸都是顺式的,但是这样的顺式会容易被氧化,但是反式不易被氧化,所以在加工过程当中可能有意或者无意在还原脂肪酸的时候并没有还原彻底,反而将顺式变为反式)
二、复合脂
复合脂即复脂,除了含有脂酰基和醇基团,还包括一些非脂成分比如磷酸基团或者是糖基。其中磷脂是含有磷酸基团的脂类,比如甘油磷脂和鞘磷脂,糖脂是含有糖基的脂类比如说甘油糖脂和鞘糖脂。其中鞘糖脂和鞘磷脂统称为鞘脂。鞘脂广泛存在于各种真核生物,但是在细菌和古菌中很罕见。
关于甘油磷脂和鞘磷脂在细胞生物学膜结构当中有详细的介绍,所以这里我不再写了哈
糖脂是糖类通过半缩醛羟基和脂质以糖苷键连接而成的糖缀合物,包括鞘糖脂和甘油糖脂,其中鞘糖脂的脂质部分是神经酰胺,甘油糖脂的脂质部分是甘油二酯。
鞘糖脂又可以分为中性鞘糖脂和酸性鞘糖脂,中性鞘糖脂的糖基没有唾液酸成分,酸性鞘糖脂的糖基有酸化的硫酸化糖基或者唾液酸。
三、异戊二烯类脂
此类脂衍生于异戊二烯,在结构上可以被剖析成若干个异戊二烯单位,主要包括萜、脂溶性维生素和类固醇。
(1)萜
萜是由若干个异戊二烯单位连接而成,连接的方式主要是头尾相连或者是尾尾相连
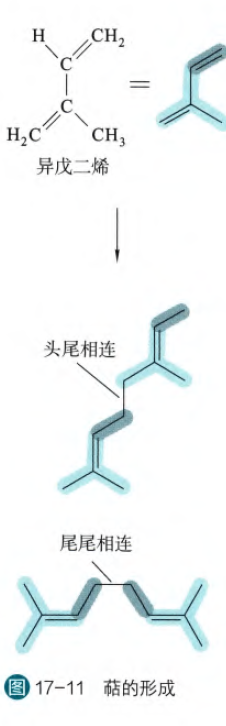
根据异戊二烯的数目,萜可以分为单萜(2个)、倍单萜(3个)、双萜(4个)、三萜(6个)、四萜(8个)等。
(2)类固醇
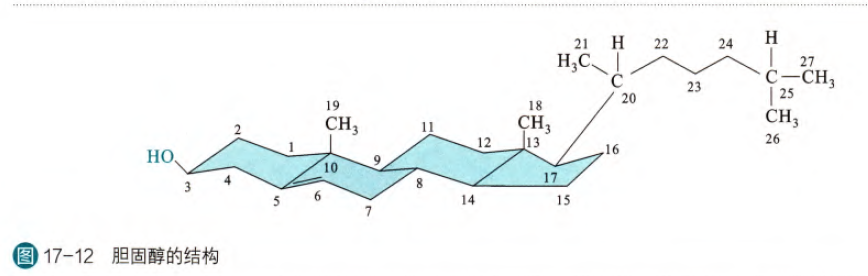
类固醇也称为甾类,其核心结构是由三个六元环和一个五元环融合而成的环戊烷多氢菲。其中胆固醇是最重要也是最常见的类固醇。胆固醇(由6个异戊二烯单位合成)的结构如下
(3)脂溶性维生素
维生素A、D、E和K都是由异戊二烯单位聚合而来,不溶于水
生物膜的结构和功能
生物膜主要是由脂质、蛋白质和糖组成的,那么糖的存在形式也大多是以糖脂和糖蛋白的形式。对于膜脂我们不再多提,可以在细胞生物学笔记当中看到,对于膜蛋白主要再强调一下他们固定(或者与膜作用的方式是什么)
膜蛋白可以分为三种:外在蛋白、内在蛋白和脂锚定蛋白
首先外在蛋白是和膜结构非常松散地结合,主要是依靠氢键或者是离子键的作用。外在蛋白可以与膜脂的亲水头部,也可以和暴露在膜表面的内在蛋白形成上述作用。因为是很松散的结合,所以膜外在蛋白通过调节pH或者高盐浓度就可以影响到氢键和离子键的作用从而脱离膜,细胞色素c就是属于线粒体内膜的外在蛋白
内在蛋白也称为整合蛋白,这种蛋白是贯穿膜的,跨膜区域主要依靠疏水的α螺旋来与膜脂分子的疏水尾部紧密结合(当然也存在β桶结构跨膜)
脂锚定蛋白不含跨膜的肽段,但是可以通过共价结合的脂质提供的疏水尾锚定到质膜的一侧。它们共价结合的脂质可以视为是蛋白质翻译后修饰的产物,这种化学修饰在一定程度上是可逆的。根据脂质的性质,膜锚定蛋白可以分为四类:
(1)第一类是通过N端的甘氨酸的氨基与脂肪酸的羧基形成酰胺键,然后通过疏水的脂酰基与膜脂的疏水核心结合锚定在质膜上,提供疏水尾的脂肪酸一般是豆蔻酸。这类脂锚定蛋白参与细胞信号转导、蛋白质与蛋白质的相互作用以及蛋白质定向。
(2)第二类是通过半胱氨酸的巯基或者是丝氨酸或者是苏氨酸的羟基与脂肪酸的羧基缩合,形成硫酯键或者酯键,然后同样通过疏水的脂酰基和膜脂的疏水核心的结合锚定在质膜上,这里提供疏水尾部的有软脂酸、硬脂酸和油酸等。这一类脂锚定蛋白主要锚定在质膜的胞内一侧,一般也参与跨膜信号的转导。
(3)第三类是通过硫醚键相连的异戊二烯化蛋白,这类脂锚定蛋白含有若干的聚合在一起的异戊二烯基团,与半胱氨酸的巯基S共价相连,通过疏水性的异戊二烯基团插入到脂双层的内部而锚定到膜上,这一类膜锚定蛋白对于真核细胞的生长、分化和形态形成非常重要。
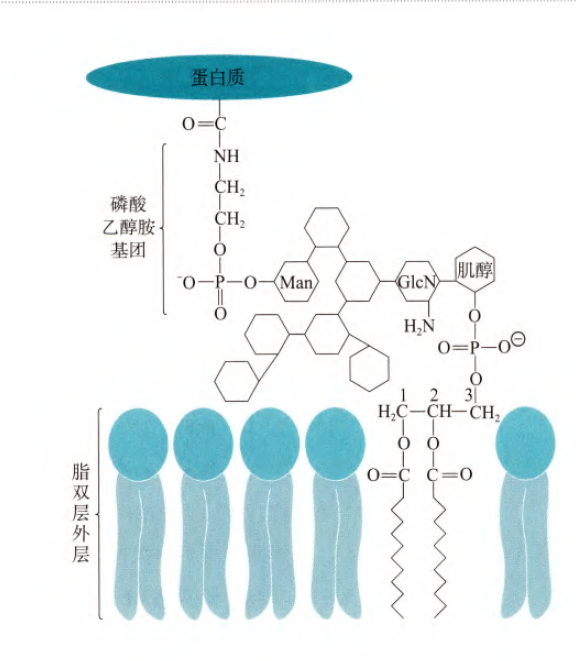
(4)最后一类是糖基磷脂酰肌醇锚定蛋白(GPI)蛋白,其C端氨基酸的羧基与磷酸化的乙醇胺相连,后者再与膜上糖基化的磷脂酰肌醇共价相连,其中寡糖链的组成和磷脂酰肌醇中脂肪酸的种类因为蛋白质的不同而有所变化(原核细胞当中目前没有发现存在GPI蛋白)(如图)
生物膜的基本结构和性质
理论上,膜脂的两性性质可以使得它们在水相当中形成四种稳定形态
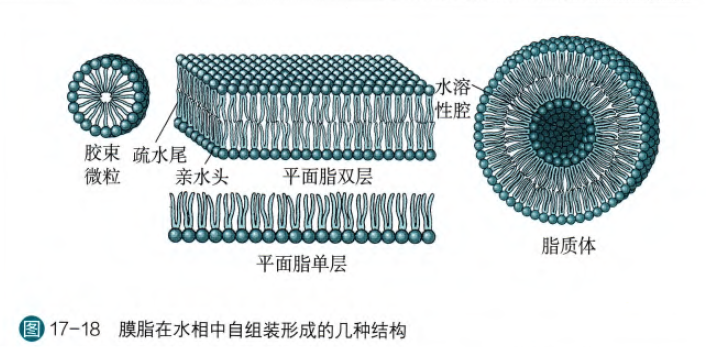
其中脂质体是最稳定的结构
关于生物膜的结构和流动性的调节在细胞生物学也有讲到,这里也不再阐述,但是有一点笔记当中没有提到的是对于细菌、真菌和植物调节膜流动性的机制,这一般是通过调节膜脂分子当中不饱和脂肪酸(不饱和程度up,流动性up)和短链脂肪酸(脂肪链越短,脂质分子间作用力越小,流动性up)的含量来调节流动性。
那么对于细菌,温度的下降可以提高在脂肪酸分子引入双键的去饱和酶的基因表达,但是对于植物和真菌而言,温度的下降实际上是通过增大氧(去饱和酶的底物之一)的溶解度从而提高去饱和酶的活性
生物膜的相变和分相
在较低温度的时候,膜脂分子的脂酰基链呈现高度有序的排列,这种有序的排列使得膜处于一种类似于固体的结构,但是随着温度的升高,膜脂上的脂酰基从有序变为无序,原先处于固态的膜脂逐渐转变为流体态或者是液晶态。生物膜这种从固态转变为液态的过程叫做相变(反过来也是),发生相变时候的温度为相变温度。根据膜脂成分的不同,在生理温度下,膜脂双分子层当中有相当一部分是流体态或者是液晶态,另外一部分表现为固体态或者结晶态,生物膜的这种性质称之为分相(生物膜的相变可能会影响到膜蛋白的功能,特别是膜蛋白的活性中心或者关键结构与膜脂相关的)