MADS-box编码基因Tunicate1通过增加玉米果穗上方的叶片数量正向调控玉米产量。

摘要
玉米果穗上方的叶片是玉米籽粒灌浆的主要碳水化合物来源。然而,在现代玉米育种中,通过增加果穗上方叶片数量来增强碳源并提高玉米产量仍然是一个挑战。在此,我们克隆了与果穗上方叶片数量相关的数量性状位点(QTL)的致因基因。该致因基因是之前报道的MADS-box结构域编码基因Tunicate1(Tu1),它负责有稃玉米或Tunicate玉米的表型。我们发现,Tu1可以在保持源—库平衡的同时显著增加果穗上方的叶片数量。源自一种爆米花地方品种的Tu1基因远端上游一个5个碱基对(bp)的插入增强了其转录,共调控其叶序激活因子和抑制因子,并增加了果穗上方的叶片数量。田间试验表明,在常规种植和密植条件下,Tu1的5-bp插入分别可以将玉米籽粒产量提高11.4%和9.5%。从地方品种中发现这种有利的Tu1等位基因表明,地方品种是玉米高产育种的宝贵资源。
引文
从有限的土地面积中优化作物产量对于应对日益增长的全球粮食需求至关重要。提高光合能力(碳源)是提高单位面积产量的有效措施。玉米的光合能力与其叶片数量和植物发育阶段密切相关。位于主果穗上方的叶片相对更年轻且代谢更活跃,而位于果穗下方的叶片则容易受到遮荫和衰老的影响。此外,果穗上方的叶片主要为果穗的籽粒灌浆提供碳水化合物,而果穗下方的叶片主要用于维持茎和根的生长。因此,玉米果穗上方的叶片数量是决定植物碳源强度、源库平衡和整体产量的重要因素。适当地增加果穗上方的叶片数量可以增强碳源强度,从而提高光合能力,并在果穗中积累更多的同化产物。这反过来会导致果穗变大和籽粒产量增加。
玉米在驯化和改良过程中,其果穗上方的叶片数量对碳源强度产生了显著变化(图1a)。野生玉米(Teosinte)通常在大多数叶腋处产生果穗。与它们的野生祖先相比,玉米地方品种通常具有更少的果穗上方叶片。相反,玉米自交系通常以果穗上方有更多的叶片为特征。因此,玉米自交系通常比野生玉米和地方品种具有更强的碳源。然而,关于玉米在驯化和改良过程中通过改变果穗上方叶片数量来增强碳源强度的遗传基础仍然知之甚少。
在胚胎发生过程中,茎尖分生组织(SAM)会早期启动,通过从叶原基依次启动形成一系列叶片,直到开花转变发生。叶片数量的产生是由叶片启动的开始时间或速率(叶序)以及营养生长阶段的持续时间决定的,而营养生长阶段的持续时间则由开花转变的时间控制。许多参与这些过程的基因已被鉴定出来。例如,拟南芥中的“暂停”(psd)突变体表现出叶片启动延迟,导致其叶片数量比野生型少。相反,玉米中的Knotted1(Kn1)基因抑制叶片启动并保持分生组织的特性。水稻中的PLASTOCHRON1(PLA1)、PLA2和PLA3基因,玉米中的TERMINAL EAR1(TE1)、WEE1和VIVIPAROUS8基因,拟南芥中的ALTERED MERISTEM PROGRAM1(AMP1)基因,以及大麦中的多节矮生基因均调控叶序,这些基因的功能缺失会导致叶片数量增加。SBP-box基因unbranched 2(ub2)调控叶片启动速率,其功能缺失会导致叶片数量增加。同样,Big embryo1(Bige1)基因参与侧生器官启动,该基因的功能缺失突变体可加速玉米叶片的启动。此外,玉米叶片数量还与MADS-box转录因子ZmMADS1和ZmMADS3、生长素外排转运蛋白PIN1a以及分生组织大小调节因子Thick Tassel Dwarf1(TD1)相关。
由于开花转变的时机是营养生长阶段持续时间的决定性因素,因此在玉米中,叶片数量常被用作开花时间的指标。然而,开花时间是作物适应性的一个关键因素,并且会影响许多其他农艺性状,因此通过延迟开花时间来增加叶片数量以提高玉米籽粒产量并不是一种合适的方法。迄今为止,尚未发现可以通过增加穗上叶片数量而不影响开花时间来提高玉米籽粒产量的可利用自然位点。
在本研究中,我们克隆了一个与穗上叶片数量相关的主要数量性状位点(QTL)的候选基因。我们发现,该基因(Tunicate1,Tu1)编码一个MADS-box蛋白。Tu1启动子上游的一个5个碱基(AGAAG)插入增强了其转录,通过调节叶序激活因子(如TD1、PIN1a和yabby1)和抑制因子(如ub2、Bige1、Kn1、ZmMADS1、ZmMADS3和WEE1),适度增加了穗上叶片数量,同时保持了大多数其他性状不变。这种有利的5个碱基插入等位基因最初来源于野生玉米。它能够在保持源库平衡的同时提高玉米产量,显示出其在玉米育种中的应用价值。
结果
Lna1 是与玉米穗上叶片数量相关的主要数量性状位点(QTL)
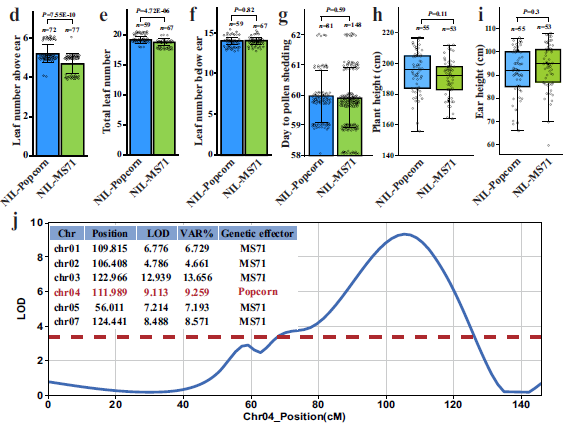
为了揭示玉米穗上叶片数量的分子遗传基础,我们将一种地方品种——草莓爆米花(以下简称爆米花)与玉米嵌套关联作图群体(NAM)的亲本自交系 MS71 进行杂交,构建了一个 F₂ 群体(图 1)。爆米花仅在穗上产生 3-4 片叶片,而 MS71 则产生 5 或 6 片叶片(图 1b)。全基因组 QTL 分析显示,在该 F₂ 群体中,位于 1、2、3、4、5 和 7 号染色体上的 6 个 QTL 与穗上叶片数量相关。在这些鉴定出的 QTL 中,位于 4 号染色体长臂上的一个 QTL 被命名为穗上叶片数量 1(Lna1),是唯一一个来自爆米花亲本的具有遗传效应的 QTL,解释了 9.3% 的总表型变异(图 1j)。
为了精确量化 Lna1 的遗传效应,我们进一步构建了一对近等基因系(NILs)(见“方法”)。与携带纯合 MS71 Lna1 等位基因的 NIL(NIL-MS71)相比,携带纯合爆米花 Lna1 等位基因的 NIL(NIL-爆米花)在穗上有更多的叶片(图 1c、d)。NIL-爆米花的总叶片数量多于 NIL-MS71,而两者的穗下叶片数量没有显著差异(图 1e、f)。穗上叶片数量的增加导致 NIL-爆米花的总叶片数量增加。尽管叶片数量通常与玉米的开花时间密切相关,但我们的结果表明,NIL-MS71 和 NIL-爆米花之间开花时间没有显著差异(图 1g)。同样,两者的株高和穗高也没有显著差异(图 1h、i)。此外,我们测量了穗上各节间的长度,发现 NIL-爆米花的第 1-4 节间(从上到下)比 NIL-MS71 更短,而第 5 节间的长度在两者之间相似(补充图 1)。这些结果表明,Lna1 可以显著增加穗上叶片数量,同时保持大多数其他性状不变。
玉米中 Lna1 位点的高分辨率定位
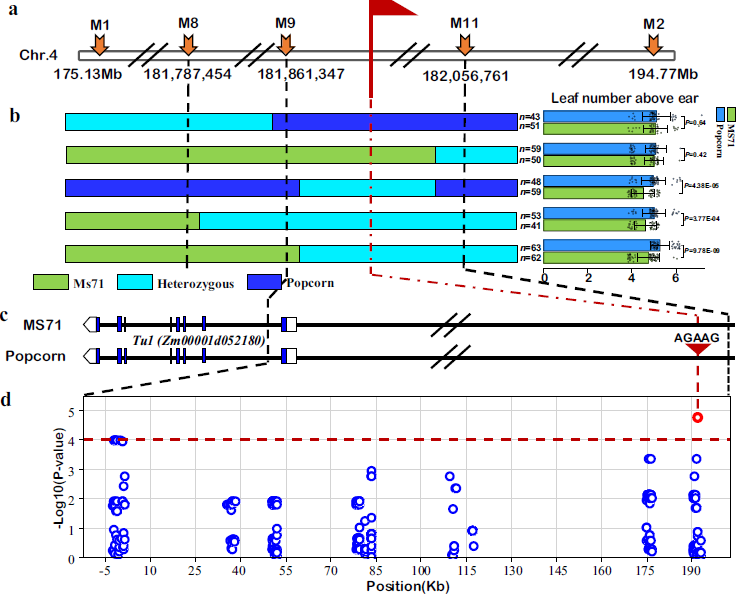
对 F₂:₃ 群体进行遗传连锁分析发现,Lna1 位于 4 号染色体长臂上的两个标记 M1 和 M2 之间(图 2a)。为了进一步精细定位 Lna1 位点,我们从 17 个代表性 F₇ 重组植株系中构建了一个包含 11914 个个体的较大群体,并采用了一种改进的后代测验策略(见方法和补充图 2)。利用 12 个简单序列重复(SSR)和插入/缺失(Indel)标记,基于 B73 基因组序列(V4;http://www.maizegdb.org),最终将 Lna1 的候选基因锁定在 181861347 至 182056761 bp 之间的一个 195 千碱基对(kb)的基因组片段内,该片段由标记 M9 和 M11 所界定(图 2b,补充图 2)。序列注释显示,该区域内仅包含一个基因(Zm00001d052180)的第一外显子(图 2c)。已知 Zm00001d052180 是导致 Tunicate1(Tu1)突变体表型(即有稃玉米或有稃玉米)的基因。因此,Zm00001d052180 被认为是 Lna1 的候选致因基因。为了避免混淆,我们在本研究中保留了之前的名称 Tunicate1(Tu1)。
Tu1 包含 8 个外显子和 7 个内含子(图 2c),编码一种含有 228 个氨基酸残基的 MADS-box 蛋白。系统发育分析显示,这种 MADS 结域蛋白属于 MIKC 型 MADS 组,并且与拟南芥的 SHORT VEGETATIVE PHASE(SVP)蛋白密切相关(补充图 3)。对 MS71 和爆米花之间的序列比较发现,在精细定位区间内,Tu1 的仅有一个外显子没有变异,但在该 195-kb 区间内,基因上游的两个亲本系之间检测到许多单核苷酸多态性(SNPs)和插入/缺失。为了确定哪个变异是导致穗上叶片数量的致因变异,我们在嵌套关联作图(NAM)群体中进行了关联作图,该群体由大约 5000 个个体组成。基于 26 个 NAM 亲本的全基因组测序数据(https://nam-genomes.org),在 195-kb 片段中共鉴定出 475 个变异。关联作图显示,与穗上叶片数量相关性最强的信号(P = 1.8×10⁻⁵)位于 Tu1 上游 -194 kb 处的远端区域的一个 5-bp 插入(AGAAG)处(图 2d)。该区域也被报道为通过转座酶可及染色质测序(ATAC-seq)数据确定的染色质可及性景观(补充图 4)。此外,该区域被鉴定为与 Tu1 远距离相互作用的 H3K27me3-HiChIP 环,而 H3K27me3 作为一种转录沉默元件(补充图 5)。由于其他变异的信号低于阈值(P = 1.0×10⁻⁴),因此与 MS71 相比,爆米花中具有最强信号的 5-bp 插入被确定为 Lna1 的致因变异。这些结果表明,Tu1 是玉米穗上叶片数量的致因基因。
通过转基因分析确认了 Tu1 基因对穗上叶片数量的影响
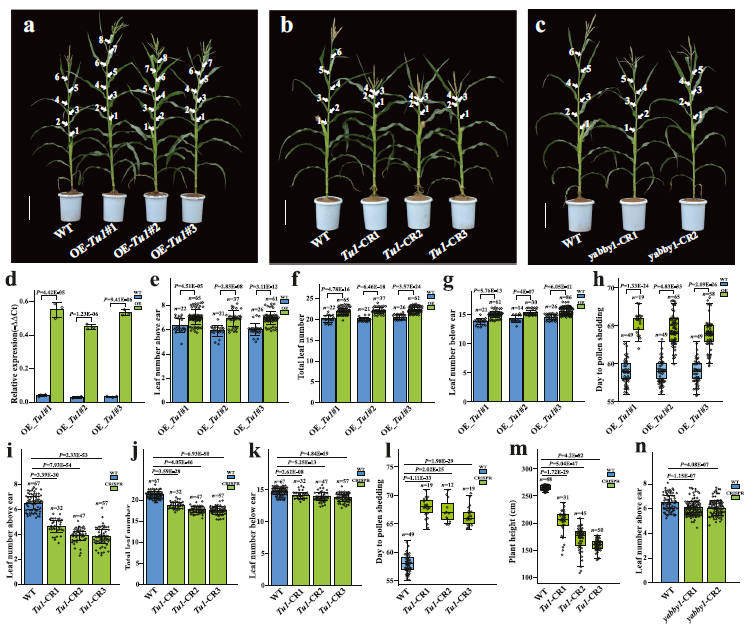
为了确定与 Lna1 对应的 Tu1(Zm00001d052180)基因在玉米穗上叶片数量中的作用,我们进行了遗传转化实验(图 3)。我们将一个由泛素启动子控制的构建(Tu1-OE)导入玉米自交系 LH244 中,成功获得了三个独立的转基因事件(图 3a)。与对照植株相比,过表达 Tu1 基因的转基因植株在穗上叶片数量和总叶片数量上显著增加(P < 2.5 × 10⁻⁵;图 3d–f)。与自然 Tu1 等位基因类似,这种 MADS-box 基因的异位过表达也在本研究中诱导了雄穗上出现大型小穗和穗上覆盖谷粒的长颖壳(补充图 6)。随后,我们利用 CRISPR/Cas9 技术对 Zm00001d052180 基因的编码序列(CDS)进行了单切割靶向玉米转化,获得了三个独立的编辑事件(Tu1-CR1、Tu1-CR2 和 Tu1-CR3),这些事件中 Tu1 基因的功能丧失(图 3b)。编辑植株在 Tu1 的第一个外显子中携带缺失突变,引入了移码突变,导致翻译提前终止(补充图 7)。这些编辑植株表现为半矮化,其穗上叶片数量显著减少(P < 2 × 10⁻³⁰),平均只有 3–5 片叶片,而对照植株平均有 6.5 片穗上叶片(图 3b,i–m)。这些转基因结果证实了 Tu1 基因(对应于 Lna1)控制玉米穗上叶片数量。
我们通过扫描电子显微镜(SEM)和光学显微镜观察了过表达和基因敲除编辑玉米植株的茎尖分生组织(SAM)中的叶序数量。在播种后 30 天(DAP),过表达植株的叶序数量为 19(补充图 8a、e、f),而基因敲除编辑植株的叶序数量为 15(补充图 8b、g、h)。到 35 DAP 时,过表达植株的叶序数量增加到最大值 21,而基因敲除编辑植株的叶序数量为 16。此时,过表达植株的 SAM 中已经出现了花序原基,而基因敲除编辑植株的 SAM 中尚未出现花序原基(补充图 8c、d)。这些结果表明,玉米的叶片数量是由 SAM 中花序原基出现之前的叶序数量决定的,且过表达植株的 SAM 活性高于基因敲除编辑植株的 SAM。
一个 5-bp 插入适度增强 Tu1 表达并略微增加穗上叶片数量
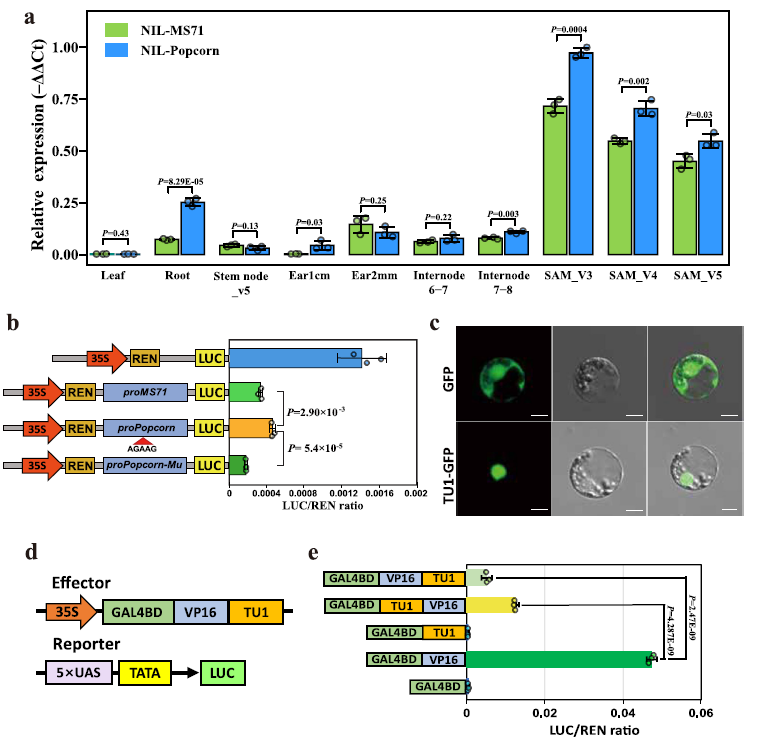
为了确定 Tu1 上游远端区域的 5-bp 插入如何调控其转录,我们在 V3-V5 阶段的 NIL-MS71 和 NIL-爆米花的茎尖分生组织(SAM)中进行了实时 RT-PCR 检测。RT-PCR 结果显示,携带 5-bp 插入的 NIL-爆米花中 Tu1 的表达水平明显高于 NIL-MS71(图 4a)。为了进一步测试这一 5-bp 插入对 Tu1 表达的影响,我们在玉米原生质体中进行了双荧光素酶(LUC)转录激活实验(图 4b)。随后,我们将含有由 MS71 和爆米花远端启动子片段驱动的 LUC 报告基因的构建转化到原生质体中。与 MS71 片段相比,爆米花片段中的荧光素酶活性更高。此外,当从爆米花片段中删除 5-bp 插入时,荧光素酶活性明显受到抑制(图 4b)。这些结果表明,5-bp 插入增强了爆米花植株中 Tu1 的表达。因此,关联作图和 LUC 瞬时表达分析一致揭示了 Tu1 基因远端启动子区域中的 5-bp 插入(AGAAG)是致因变异。
为了阐明 Tu1 在决定穗上叶片数量中的调控作用,对与叶片数量相关的性状进行全面调查是必要的,这些性状包括穗上/下叶片数量、开花时间(DTP)以及穗出现的节点(相当于穗下叶片数量)(图 1d–i 和图 3e–l)。将这些性状在过表达和编辑转基因植株与非转基因对照植株之间进行比较,揭示了稳健的模式。过表达转基因植株的穗上和穗下叶片数量显著增加,从而导致总叶片数量大幅增加(图 3e–g)。这些过表达植株的穗出现节点平均比对照植株多一个节间(图 3g)。此外,与对照植株相比,过表达植株的平均开花时间显著延迟(图 3h)。
相反,经过基因编辑的转基因植株表现出穗上和穗下叶片数量的显著减少,从而导致总叶片数量的大幅降低(图 3i–k)。与对照植株相比,编辑植株的穗出现节点平均减少了接近一个节间(图 3k)。然而,编辑植株的平均开花时间(DTP)显著延迟(图 3l),与过表达植株相似。这些发现突出了 Tu1 在控制穗上和穗下叶片数量中的作用,它影响了转基因植株的穗出现节点和开花时间。相比之下,携带 Tu1 启动子中 5-bp 插入的植株,其 Tu1 表达适度增加,调节总叶片数量而不影响穗出现的节点和开花时间(图 1f,g)。因此,带有 5-bp 插入的植株的穗下叶片数量没有差异,而穗上叶片数量受到调控(图 1d,f)。过表达植株(表达增强)和编辑敲除植株(表达过度抑制),以及带有 5-bp 插入的植株,一致表明 Tu1 调控总叶片数量并无矛盾。与过表达和过度抑制植株不同,带有 5-bp 插入且 Tu1 表达适度增强的植株并未改变穗出现的节点或开花时间,这表明这些因素的变化取决于 Tu1 的表达水平。因此,Tu1 对穗上叶片数量的调控归因于穗出现节点和开花时间的不变,这是由于带有 Tu1 启动子 5-bp 插入的玉米植株中 Tu1 表达适度增加的结果。
TU1 可能作为转录抑制因子发挥作用
Tu1 基因在叶片、茎节、节间、根以及 1 厘米或 2 毫米长的幼穗中的表达水平较低,但在茎尖分生组织(SAM)中显著更高(图 4a)。从 V3 到 V5 发育阶段,Tu1 转录本在 SAM 中的积累逐渐减少,无论发育阶段如何,NIL-爆米花中的转录本丰度均高于 NIL-MS71(图 4a)。
TU1 蛋白是一种 MADS-box 转录因子(补充图 3)。为了确定 TU1 的亚细胞定位,我们将编码 TU1-GFP 融合蛋白的构建导入玉米叶片原生质体中,结果显示 TU1-GFP 的荧光信号仅在细胞核中被检测到,而游离 GFP 则没有(图 4c)。为了检测 TU1 的自我激活活性,我们进行了酵母双杂交实验,结果表明 TU1 和 BD(GAL4 DNA 结合域)融合蛋白并未激活报告基因的表达,这表明 TU1 缺乏转录激活活性(补充图 9)。为了进一步确认这一点,我们通过构建嵌合蛋白进行了转录活性实验,其中 TU1 与酵母 GAL4 转录因子的 DNA 结合域(GAL4-DB)和单纯疱疹病毒蛋白 16 的激活域(VP16)融合(图 4d)。这些实验的报告构建包含由合成启动子驱动的荧光素酶报告基因,该合成启动子包含五个 GAL4 上游激活序列(UAS)拷贝和一个 TATA 盒(图 4d)。虽然 GAL4BD-VP16 强烈激活了荧光素酶表达(表现为高荧光素酶活性),但 GAL4BD-VP16-TU1 嵌合蛋白显著抑制了同一报告基因的荧光素酶活性(图 4e)。这些结果表明,TU1 可能作为其下游基因的转录抑制因子发挥作用。
我们重新分析了一个先前报道的玉米单细胞 RNA 测序数据集,重点关注茎尖分生组织(SAM),并遵循已报道的方法。结果显示,Tu1 在 20 种不同的细胞类型中均有表达,且在叶原基细胞中表现出明显的表达(补充图 12),这与基于原位杂交的转录模式一致(补充图 8i)。在选定的 9 个下游目标基因中,除了在该单细胞 RNA 测序数据集中未检测到的 ZmMADS3 外,我们在叶原基细胞中识别出与这些基因相关的表达模式,为它们与 TU1 在调控叶片数量中的关联提供了有力证据。
接下来,我们对 TU1 进行了 DNA 亲和纯化测序(DAP-seq)(补充数据 4)。分析发现,TU1 在 20821 个基因的调控区域中有 52806 个结合峰,其中“GG(A/T)₆CC”这一基序在这些结合位点中高度富集(图 5b)。这一基序与先前鉴定的 MADS-box 转录因子的结合位点一致。ZmMADS1 基因的启动子区域含有 TU1 的结合位点“GG(A/T)₆CC”(图 5c)。ZmMADS1 基因的表达水平受到 TU1 的抑制。此外,ZmMADS1 启动子中包含“GG(A/T)₆CC”基序的区域显示出较高的染色质可及性(补充图 13)。荧光素酶活性实验和电泳迁移率变化实验(EMSAs)证实,TU1 可以直接结合到 ZmMADS1 启动子中的这些基序,并随后抑制其表达(图 5d-f)。
这些结果表明,TU1 可以直接结合到 ZmMADS1 的启动子上,从而直接抑制其表达。此外,TU1 间接上调了 TD1、ZmMADS3、PIN1a、Kn1、Bige1、ub2 和 yabby1 的表达,并间接下调了 WEE1 的表达(图 5g)。其中,TD1(补充图 14)、PIN1a 和 yabby1 对叶片数量有积极影响,而 WEE1、ZmMADS1、ZmMADS3、Kn1、ub2 和 Bige1 对这一性状有负面影响(图 5g)。因此,TU1 调控这些基因的作用,从而温和地增加了玉米的总叶片数量。
Lna1 位点在玉米多样化过程中受到选择
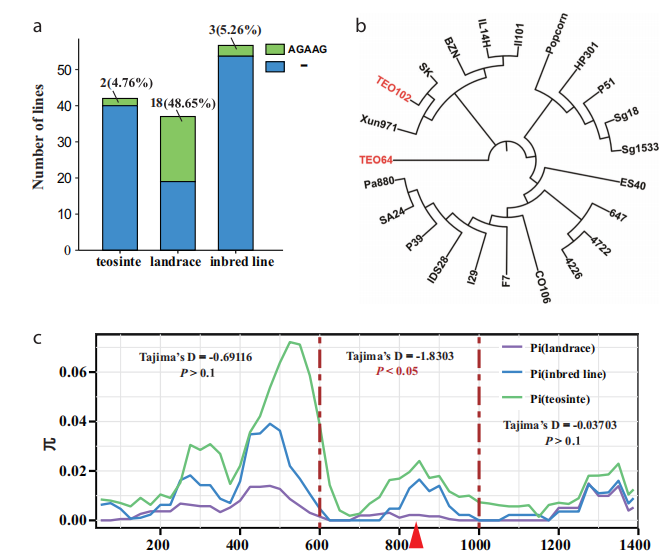
为了研究 Lna1 位点是否在玉米中受到选择,我们对来自 136 份材料(包括 42 份野生玉米、37 份地方品种和 57 份普通玉米自交系)的 1.4-kb 启动子片段(包含一个 5-bp 插入/缺失)进行了大规模测序(补充数据 5)。大多数材料中不存在 5-bp 插入,除了 4.76%(2 份)的野生玉米、48.65%(18 份)的地方品种和 5.26%(3 份)的玉米自交系(图 6a)。这一结果表明,爆米花的 Lna1 等位基因在野生玉米向地方品种驯化过程中积累,但在玉米改良过程中逐渐丢失。此外,我们对所有携带 5-bp 插入的材料(23 份)进行了系统发育分析,结果表明,携带 5-bp 插入(AGAAG)的野生玉米和玉米材料共享一个共同祖先(图 6b)。随后,我们比较了野生玉米、地方品种和玉米自交系之间的核苷酸多样性。野生玉米表现出最高的 DNA 多样性,而玉米自交系的多样性较低,地方品种的多样性最低。我们将 1.4-kb 片段分为三个片段(1–600 bp、601–1000 bp 和 1001–1400 bp)。随后,我们对这些材料的三个片段进行了 Tajima’s D 测试。Tajima’s D 测试仅在地方品种的第二个片段(601–1000 bp,包含 5-bp 插入)中显著拒绝了中性假设(Tajima’s D = -1.8303,P < 0.05),而在野生玉米或玉米自交系的任何测序区域中均未检测到显著的 Tajima’s D 信号(图 6c)。这些发现表明,Lna1 位点在玉米多样化过程中受到选择。
爆米花的 Lna1 等位基因通过增加穗上叶片数量提高玉米产量
序列分析结果显示,在 42 份野生玉米中仅有 2 份,而在 94 份玉米材料中有 21 份携带 5-bp 插入。大多数携带该插入的玉米材料是地方品种。这些发现表明,5-bp 插入在野生玉米中是一种罕见变异,主要在地方品种中积累,但在玉米改良过程中大部分已经丢失。
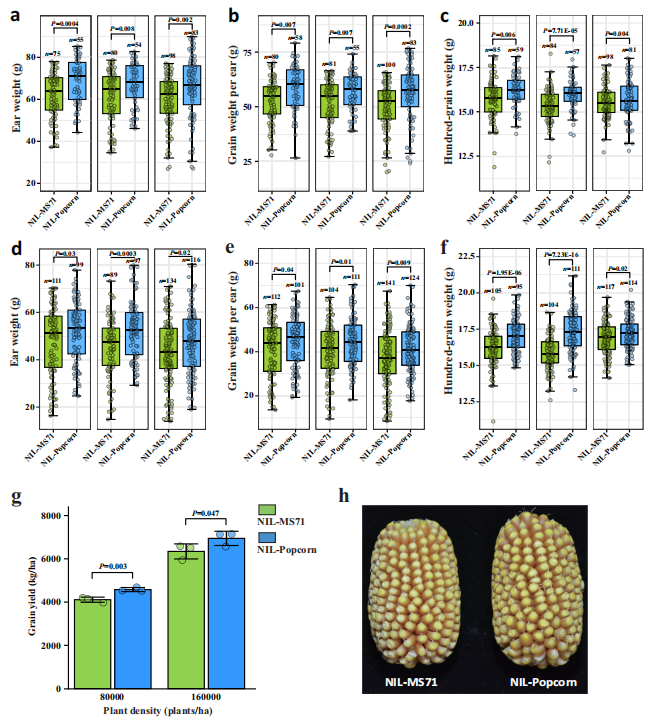
尽管在玉米改良过程中几乎完全丢失,但已知可以增加穗上叶片数量的 Lna1 位点中的 5-bp 变异,有可能被用于优化株型,特别是在现代育种计划中的密植条件下。为了验证这种可能性,我们在正常密度(每公顷 8000 株,图 7a–c)和高密度(每公顷 16000 株,图 7d–f)条件下种植了三组 NIL-MS71 和 NIL-爆米花品系,并进行了产量测试(见“方法”)。在正常种植密度下的结果显示,与 NIL-MS71 相比,NIL-爆米花的单穗重量、穗粒重和百粒重分别平均提高了 10.2%、11.4% 和 2.6%。当 NIL 品系以双倍密度种植时,NIL-爆米花的单穗重量、穗粒重和百粒重分别平均提高了 10.8%、9.5% 和 5.5%。这些结果表明,爆米花的 Lna1 等位基因可以在正常密度和高密度田间条件下增加穗上叶片数量和产量潜力(图 7g,h)。因此,爆米花的 Lna1 等位基因可用于改善株型,并在常规种植和密植田间提高玉米产量。