芍药BAHD酰基转移酶-文献精读128
PoDPBT, a BAHD acyltransferase, catalyses the benzoylation in paeoniflorin biosynthesis in Paeonia ostii
PoDPBT,一种BAHD酰基转移酶,在芍药(Paeonia ostii)中催化芍药苷生物合成中的苯甲酰化反应。

摘要
PoDPBT是属于BAHD家族的一种O-苯甲酰基转移酶,能够催化8-去苯甲酰基芍药苷转化为芍药苷。PoDPBT是第一个被证实参与芍药苷生物合成修改阶段的酶。在BAHD家族中发现了一个新的DFGGG模式,类似于DFGWG。转录组数据库为进一步研究参与芍药苷生物合成的其他酶基因提供了资源。
关键词:O-苯甲酰基转移酶,芍药苷生物合成,单萜,芍药(Paeonia ostii),BAHD酰基转移酶
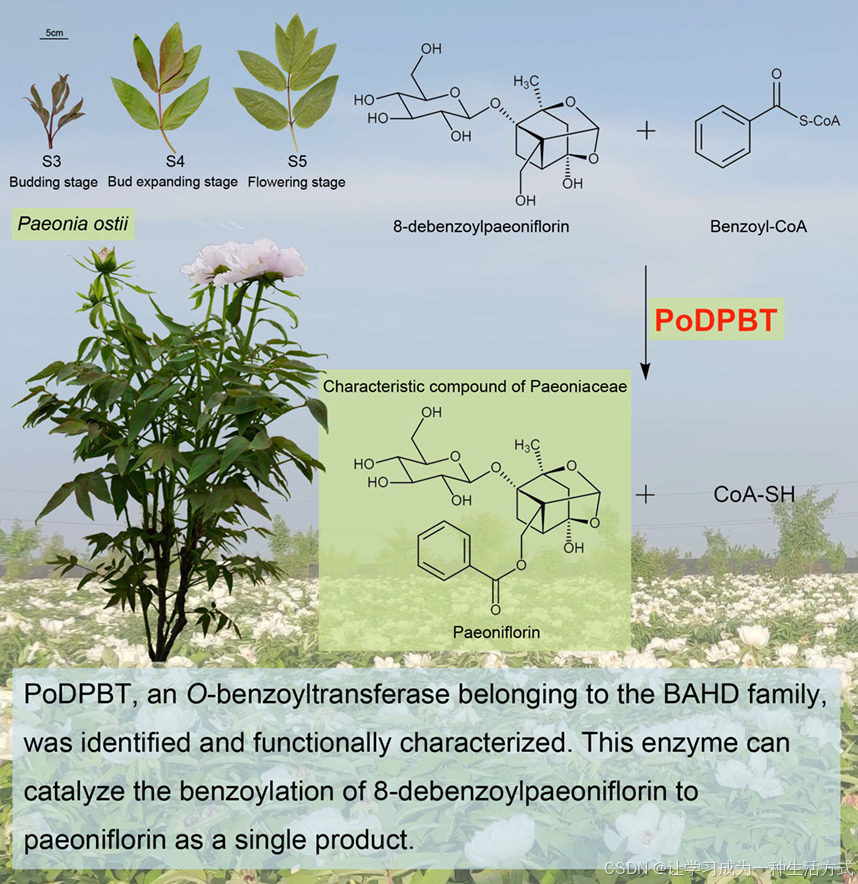
芍药苷是一种双环单萜糖苷,是一种具有多种药理活性的重要药用化合物,如抗抑郁、抗血栓、免疫调节和神经保护等(张等人,2022年;赵等人,2020年)。此外,芍药苷仅存在于芍药科植物中,被认为是芍药科的特征性化合物(张等人,2019年)。然而,其生物合成途径仍需完全阐明。苯甲酰辅酶A:苯甲醇O-苯甲酰基转移酶(BEBTs)属于BAHD酰基转移酶家族,这些酶对苯甲酰辅酶A作为酰基供体具有高亲和力,可以利用不同的芳香醇作为酰基受体(Chedgy等人,2015年)。8-去苯甲酰基芍药苷是芍药苷的衍生物之一,也广泛存在于芍药科植物中。与芍药苷不同,8-去苯甲酰基芍药苷在C8位置具有羟基而非苯甲酰基,因此其结构类似于芳香醇。因此,我们推测芍药科植物中存在的一种O-苯甲酰基转移酶可以利用苯甲酰辅酶A和8-去苯甲酰基芍药苷作为底物来合成芍药苷(张等人,2022年)。这种酶将被指定为苯甲酰辅酶A:8-去苯甲酰基芍药苷8-O-苯甲酰基转移酶(DPBT)。
我们之前发现牡丹叶中的芍药苷含量与植物发育阶段显著相关(张等人,2019年)。为了筛选参与芍药苷生物合成的O-苯甲酰基转移酶编码基因,分别在牡丹发育的萌芽阶段(S3)、芽扩张阶段(S4)和开花阶段(S5)收集叶片的RNA样本(图S1),然后在BGISEQ-500平台上进行测序(图1a)。通过qRT-PCR验证了RNA-Seq数据的可靠性(图S2和表S1)。基于基因注释,确定了23个O-苯甲酰基转移酶候选基因(表S2),这些基因在不同发育阶段表现出不同的表达模式(图1b)。其中,Pos.gene26002、Pos.gene30573、Pos.gene35667、Pos.gene79215和Pos.gene81370的表达水平与芍药苷含量高度相关(相关系数>0.9)。然而,除了Pos.gene30573外,其他四个基因在所有三个发育阶段的FPKM值都低于1.00(表S2)。因此,我们关注Pos.gene30573在将8-去苯甲酰基芍药苷转化为芍药苷的作用,并将其命名为PoDPBT。
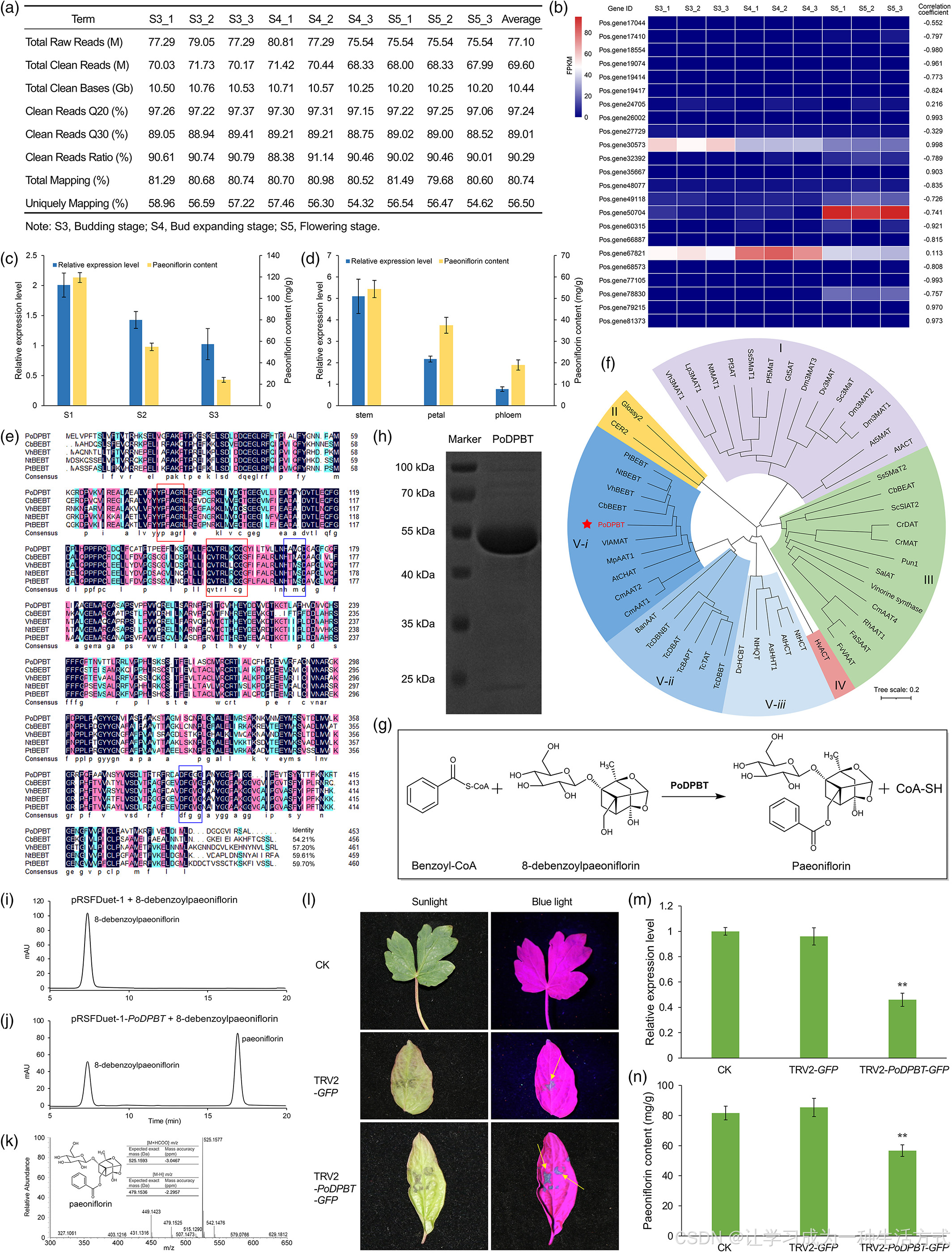
PoDPBT催化芍药苷生物合成中的苯甲酰化。 (a) RNA-Seq原始数据信息。(b) 牡丹三个发育阶段叶片中23个苯甲酰基转移酶候选基因的表达水平及其与芍药苷含量的相关性。(c, d) 不同发育阶段(c)和器官(d)中PoDPBT的表达水平和芍药苷含量。(e) PoDPBT与BEBTs的多序列比对。蓝色框显示HXXXD和DFGGG基序。红色框显示YPLAGR和QVTRLKCGG基序。(f) PoDPBT和已鉴定的BAHD成员的邻接连接法系统发育树。(g) PoDPBT催化的反应。(h) 纯化蛋白的SDS-PAGE分析。(i-k) 通过体外实验对PoDPBT的表征。空载体酶与8-去苯甲酰基芍药苷反应生成的产物的UPLC谱图(i)和PoDPBT与8-去苯甲酰基芍药苷反应产物的UPLC谱图(j);PoDPBT酶实验产物的质谱数据(k)。(l-n) PoDPBT对芍药苷产量的沉默效应。在蓝光下,TRV2-GFP和TRV2-PoDPBT-GFP感染的牡丹叶片与正常植物对照(CK)的图像,6天后(l);不同处理叶片中PoDPBT的表达水平,通过qRT-PCR测定(m);不同处理叶片中的芍药苷含量(n)。星号表示统计学上的显著差异(**P < 0.01,学生t检验)。
我们随后通过qRT-PCR检测了牡丹不同叶片发育阶段中PoDPBT的表达模式。在S3阶段,PoDPBT的表达量最高,其次是S4和S5阶段,这与芍药苷的积累一致(图1c)。我们之前的研究表明,芍药苷的水平在不同的牡丹器官中有所不同(张等人,2019年)。我们也检查了表达模式,结果显示PoDPBT在含有最多芍药苷的茎中表达量最高(图1d)。这些结果进一步确认了PoDPBT的表达水平与芍药苷含量之间的相关性,表明PoDPBT参与芍药苷的生物合成。
序列分析显示,PoDPBT编码一个预测的453氨基酸蛋白,分子量为49.91 kD。PoDPBT含有对BAHD成员高度保守的HXXXD基序,以及另一个保守区域DFGWG基序,其中色氨酸被甘氨酸替代。除了这两个保守基序外,还识别出了YPLAGR和QVTRLKCGG基序(图1e)(Tuominen等人,2011)。这些结果表明PoDPBT是BAHD酰基转移酶家族的成员。系统发育树显示PoDPBT归类于BAHD类群V-i(图1f),此类群的成员催化形成挥发性和非挥发性酯。PoDPBT与CbBEBT、NtBEBT(D'Auria等人,2002)、VhBEBT(Togami等人,2006)和PtBEBT(Chedgy等人,2015)关系密切。这些结果表明PoDPBT可以利用苯甲酰辅酶A和8-去苯甲酰基芍药苷合成芍药苷(图1g)。
PoDPBT在大肠杆菌中表达,纯化的蛋白大小与预期的49.91 kD一致(图1h)。使用苯甲酰辅酶A和8-去苯甲酰基芍药苷进行了体外酶活性测定。UPLC-MS结果显示,实验样品中的新峰与真正的芍药苷标准的保留时间和MS光谱相同(图1i-k)。这些结果证实PoDPBT作为一个O-苯甲酰基转移酶,能将8-去苯甲酰基芍药苷转化为芍药苷作为单一产物。为了确定其亲和力,使用不同浓度的8-去苯甲酰基芍药苷进行动力学分析,PoDPBT展示了米氏常数(Km)值为1364.54±166.28 μm,估算的催化常数(Kcat)值为0.54±0.05 s−1。
为了进一步验证PoDPBT在体内的功能,我们使用病毒诱导的基因沉默系统来抑制牡丹叶片中PoDPBT的表达。构建了一个含有绿色荧光蛋白(GFP)作为报告基因的烟草缩叶病毒2(TRV2)载体。在用TRV2-GFP和TRV2-PoDPBT-GFP浸润的叶片中,在蓝光下观察到绿色荧光斑点(图1l),但在正常植物中没有观察到。与正常植物和TRV2-GFP系列相比,用TRV2-PoDPBT-GFP浸润的叶片中PoDPBT的表达水平显著降低,后两者之间没有显著差异(图1m)。TRV2-PoDPBT-GFP系列叶片中检测到的芍药苷含量也最低(图1n),剩余的芍药苷是由于剩余的PoDPBT产生的。这些结果进一步证实了PoDPBT在芍药苷生物合成中的作用。
总的来说,PoDPBT是属于BAHD家族的一个O-苯甲酰基转移酶,已被鉴定并功能性地表征,该酶能将8-去苯甲酰基芍药苷催化为芍药苷作为单一产物。PoDPBT是参与芍药苷生物合成修改阶段的第一个酶。已预测此阶段涉及的其他酶,如羟化酶和糖基转移酶(张等人,2022年),这些酶将根据本研究建立的研究策略陆续进行表征。这些结果将为完全阐明芍药苷生物合成途径奠定基础。
方法
RNA测序和生物信息学分析
从芽生长阶段(S3)、芽扩张阶段(S4)和开花阶段(S5)的叶片中分别提取总RNA,使用的是RNA Prep Pure Plant Plus试剂盒(天根生物科技有限公司,北京,中国)。分别使用NanoDrop 2000(赛默飞世尔科技公司,加利福尼亚州,美国)和Agilent Bioanalyzer 2100(安捷伦科技公司,加利福尼亚州,美国)评估RNA的质量和数量。
图书馆构建由北京基因组研究所(武汉,中国)执行。mRNA通过Oligo (dT)附着的磁珠富集。目标RNA用片段缓冲液分割,然后通过随机六聚体引物引导的逆转录生成双链cDNA(dscDNA)。dscDNA在5'端经过磷酸修复,在3'端加上粘性“A”,然后在3'端粘上带有粘性“T”的接头。连接产物通过两个特异性PCR引物进行扩增。随后,PCR产物通过热变性,单链DNA通过夹板寡核苷酸和DNA连接酶环化。最终,cDNA文库在BGISEQ-500平台上进行测序。
所有原始测序读数通过SOAPnuke(李等,2008)过滤以去除低质量读数(接头序列,含有超过5%不明‘N’碱基的读数,以及含有超过20%低质量碱基的读数)。清洁读数以FASTQ格式存储,并通过HISAT2(金等,2015)和Bowtie2(Langmead和Salzberg,2012)映射到牡丹基因组序列。基因表达水平通过RSEM(李和Dewey,2011)计算,并以FPKM值(Mortazavi等,2008)表示。差异表达分析使用DESeq2(Love等,2014)进行,Q值≤ 0.05。为了识别基因的功能注释,对蛋白质数据库进行了BLASTx搜索(E值 < 0.00001),包括非冗余蛋白序列(Nr)数据库和京都基因与基因组百科全书(KEGG)。基于NR注释,使用Blast2GO程序(Conesa等,2005)(E值 < 0.00001)获取基因本体(GO)注释。
qRT-PCR分析
如上所述,从叶片、茎、花瓣和根部提取总RNA。每个样本使用1 μg的总RNA,通过带有gDNase的FastKing RT试剂盒(天根,北京,中国)制备一链cDNA。PCR在ABI StepOnePlus® 实时PCR系统(应用生物系统公司,加利福尼亚州,美国)上进行,按照制造商的说明进行。每个反应混合物(20 μl)包含10 μl的TB Green Premix Ex Taq II(Tli RNaseH Plus)(Takara),每个引物0.8 μl(10 μM),cDNA模板0.3 μl(1 μg)和8.1 μl的无酶RNase水。每个基因的PCR均进行三次重复,热循环条件如下:95°C 30秒;40个循环,95°C 5秒和64°C 30秒;95°C 15秒。通过熔解曲线分析确认引物特异性。测试基因的相对表达水平通过2-ΔΔCt方法计算,使用Actin(Pos.gene4533)基因作为内部对照。数据为三个生物重复的平均值±标准偏差。本研究中使用的基因特异性引物列在表S3中。
系统发育分析
使用DNAMAN 7(Lynnon Biosoft,魁北克,加拿大)进行多序列比对。使用MEGA X软件(Kumar等,2018)生成邻接连接树,设置选项如下:1000次bootstrap复制,Poisson模型,均匀率和成对删除。树中使用的蛋白质的GenBank登录号:Vh3MAT1, AAS77402; Lp3MAT1, AAS77404; 等等。
蛋白表达和纯化
从牡丹参考基因组获得了PoDPBT的全长序列。使用特定的正向和反向引物扩增PoDPBT的编码区。PCR产物经BamH I和Not I酶切后,克隆到带有六组氨酸-SUMO标签的pRSFDuet-1载体中。蛋白表达和纯化按照先前描述的方法进行(李等,2017)。使用SDS-PAGE分析鉴定纯化蛋白,并将其存储在含有20 mM Tris-pH 7.0、500 mM NaCl和5 mM二硫苏糖醇的缓冲液中。使用Bradford方法确定浓度,以BSA作为标准(Bradford,1976)。
酶测定和动力学
体外酶测定在一个40 μL的反应体系中进行,包括5 μL 8-去苯甲酰基芍药苷(20 mM)、5 μL苯甲酰辅酶A(50 mM)和30 μL纯化蛋白(0.64 mg/ml)。反应在30℃下进行1小时,通过液氮急速冷冻停止。用160 μL甲醇提取反应产物并超声波处理1分钟。混合物通过0.22毫米过滤器过滤,然后通过超高效液相色谱-质谱联用仪(UPLC-MS)进行分析。空载体酶作为阴性对照。
为了进行动力学测量,将0.5至20 mM范围内的8-去苯甲酰基芍药苷添加到上述反应体系中。反应时间为5分钟。米氏常数(Km)和催化常数(Kcat)值使用GraphPad Prism 7软件(GraphPad Software, Inc.)评估。数据为三个生物重复的平均值±标准偏差。
UPLC-MS分析
色谱分析使用Thermo Scientific Syncronis C18柱(100 mm × 2.1 mm,1.7 µm),基于Dionex UltiMate 3000 UPLC系统(Thermo Fisher Scientific,马萨诸塞州,美国)进行。质谱采集使用Q Exactive Plus杂交四极杆-轨道阱质谱仪(Thermo Fisher Scientific,马萨诸塞州,美国)。详细参数在我们之前的研究中有描述(张等人,2019)。使用真正的芍药苷绘制标准曲线进行定量。所有样本均进行三次重复测试,每个样本注射一次。数据为平均值 ± 标准偏差。
病毒诱导的基因沉默分析
如之前描述(谢等人,2019;杨等人,2021),通过病毒诱导的基因沉默在叶片中沉默PoDPBT。构建了一个带有绿色荧光蛋白(GFP)作为报告基因的烟草缩叶病毒2(TRV2)载体。TRV1和TRV2-GFP质粒保存在我们的实验室中。为了生成TRV2-PoDPBT-GFP构建,使用特定的引物(表S3)通过PCR扩增了332-bp的PoDPBT片段。所有三个TRV构建被转化到农杆菌株GV3101中,然后在28°C下连续振荡培养220 r/min过夜。细菌细胞在4000 g下离心并在渗透缓冲液(10 mM MgCl2、10 mM MES和200 μM 醋酰梧桐素)中重新悬浮,最终OD600为1.0。含有TRV1和TRV2构建的培养物在28°C下轻轻摇动6小时并以1:1体积比混合。对于真空渗透,整个植物在0.8 MPa压力下浸泡在渗透缓冲液中20分钟。正常植物的叶片和用TRV2-GFP浸润的植物叶片用作对照。渗透后6天,收集叶片进行GFP分析。渗透后10天,收集叶片进行qRT-PCR和芍药苷定量。数据为三个生物重复的平均值 ± 标准偏差。